ARTÍCULOS IV CONGRESO COLOMBIANO DE BOTÁNICA
EFECTO DEL GRADIENTE ALTITUDINAL SOBRE ASPECTOS AUTOECOLÓGICOS DE ESPELETIA PYCNOPHYLLA ssp. ANGELENSIS CUATREC. (ASTERACEAE) EN EL PÁRAMO EL INFIERNILLO (NARIÑO–COLOMBIA)
THE EFFECT OF THE ALTITUDINAL GRADIENT ON AUTOECOLOGICAL FEATURES OF ESPELETIA PYCNOPHYLLA ssp. ANGELENSIS CUATREC. (ASTERACEAE) IN THE PARAMO 'EL INFIERNILLO' (NARIÑO–COLOMBIA)
Iván F. Benavides–Martínez1, Diana L. Burbano–Martínez2, Sandra M. Urbano–Apráez3, María E. Solarte–Cruz4
]]> 1 Grupo de Investigación Biología de Páramos y Ecosistemas Andinos. Departamento de Biología, Sistema de Investigaciones de la Universidad de Nariño. A. A. 117576. San Juan de Pasto (Nariño), Colombia. pipeben@mail.udenar.edu.co.
Resumen
Se estudió el efecto de la elevación sobre algunos aspectos autoecológicos de la especie Espeletia pycnophylla ssp. angelensis Cuatrec. (Asteraceae), tales como su estructura demográfica, densidad poblacional, patrón de distribución espacial, producción de estructuras reproductivas, morfometría de adultos y supervivencia, en un páramo seco del departamento de Nariño (Colombia). Las hipótesis propuestas por otros autores acerca del efecto lineal que la elevación produce sobre estas variables, fueron completamente rechazadas. La elevación induce un efecto coenoclínico sobre la densidad poblacional y ésta a su vez, se correlaciona negativamente con la producción de estructuras reproductivas y el tamaño de los adultos, evidenciándose un efecto inequívoco del fenómeno de la densodependencia sobre el potencial biótico de la población. En la zona intermedia del gradiente, la densidad poblacional es máxima, pero los tamaños corporales son reducidos, la producción de estructuras reproductivas es mínima y la probabilidad de que un juvenil alcance edades avanzadas es baja. Evidentemente, esta elevación intermedia estaría presentando condiciones ecológicas favorables para el establecimiento y supervivencia de plántulas, lo que a largo plazo conlleva a un aumento de la densidad poblacional. Se propone que las generalizaciones de otros autores acerca de la influencia de la elevación sobre la autoecología de estas plantas son inverosímiles y limitadas, ya que la historia particular de vida y disturbio del páramo y los aislamientos espaciales, ecológicos y genéticos, son factores explicativos que no pueden aislarse del contexto. Paralelamente, la población se ajustó al modelo demográfico logarítmico, revelando su temprano estado de regeneración frente a la última quema ocurrida en el 2003, y la similitud de las curvas de supervivencia entre todas las elevaciones, probó una alta estabilidad demográfica.
Palabras clave: autoecología, Espeletia pycnophylla ssp. angelensis, gradiente altitudinal, Nariño, páramo
Abstract
Some autoecological features of Espeletia pycnophylla ssp. angelensis Cuatrec. such as its demographic structure, population density, spatial pattern of dispersion, production of reproductive structures, adult morphometry and survivorship, were studied in an altitudinal gradient on a dry paramo of Nariño (Colombia). Hypothesis proposed by other authors about a linear effect caused by the elevation on these variables were completely rejected. The elevation induces an ecoclinal effect on population density, and this density correlates negatively with both production of reproductive structures and adult sizes. These results elucidate an unequivocal effect of density–dependence phenomena on population fitness. The intermediate zone of the altitudinal gradient exhibits a high population density, but on the other hand show reduced corporal adult sizes, minimal production of reproductive structures and the probability of juveniles reaching advanced ages is low. Evidently, this intermediate elevation can have favorable ecological conditions for the seedling establishment, wich increments population densities at long term. We propose that other author's generalizations about the influence of elevation on the autoecology of these plants are limited, because the particular life and disturbance stories of paramos and the spatial, ecological and genetic isolating are explicative factors impossible to take away from the context. The demographic structure of the population fitted the logarithmic model, revealing its early regeneration state from the last fire event on 2003 and the similarity of the survivorship curves among every elevation, probed a high demographic stability.
Key words: autoecology, Espeletia pycnophylla ssp. angelensis, altitudinal gradient, Nariño, paramo
]]>
INTRODUCCIÓN
Smith (1980), en sus trabajos con Espeletia schultzii Weed., (Asteraceae) propuso uno de los modelos más conocidos y generalizados acerca del efecto del gradiente altitudinal sobre la densidad y el tamaño de las rosetas gigantes del género Espeletia en los páramos suramericanos. Según su hipótesis, la elevación produce un efecto lineal negativo sobre la densidad de individuos juveniles, haciendo que en las zonas subparamunas la probabilidad de que una plántula se establezca sea alta, pero disminuya de forma proporcional al aumentar la elevación. Esto se explica por que la temperatura, la tasa de descomposición orgánica y la disponibilidad de nutrientes se reducen con la elevación, perjudicando el establecimiento de plántulas. Este fenómeno tiene un efecto relacionado con lo que Smith (1980) denominó competencia intraespecífica inter–edades, que explica que plantas de diferentes edades compiten por recursos y espacio. De esta forma, en una zona de densidad poblacional elevada, la competencia es intensa y ocasiona que los individuos no alcancen edades avanzadas ni grandes tamaños corporales, restringiendo las plantas más altas y longevas cerca al superpáramo, donde la densidad poblacional y la competencia inter–edades son reducidas. Sinópticamente, al aumentar la elevación disminuye la densidad poblacional y aumenta la altura corporal promedio de los individuos adultos. Sin embargo, hay otros factores sinérgicos que fortalecen este patrón, como por ejemplo la disminución de pestes, herbivoría y daños antropogénicos a mayores elevaciones. Otros estudios (Meinzer et al., 1985; Smith, 1981) han reafirmado este patrón para la misma especie.
Smith (1980) generalizó éste modelo para todo el género Espeletia, pero propuso tres excepciones acerca de especies que no lo cumplen, dentro de las que incluyó a E. pycnophylla Cuatrec. Según sus observaciones en el páramo del volcán Puracé en Colombia, era evidente que esta especie discrepaba del patrón general, e incluso exhibía una relación contraria: plantas altas en el subpáramo y pequeñas a mayores elevaciones. Propuso entonces que en esta especie –cuya distribución se da principalmente en páramos secos– la altura corporal está determinada por la humedad edáfica, y los individuos de mayor altura corporal se encuentran en el subpáramo, donde hay un aporte hídrico permanente desde el bosque altoandino. La humedad disminuye con la elevación, y con ésta, la altura promedio de los individuos adultos.
Kovář (2001), estudiando el efecto de las quemas sobre la densidad poblacional, tamaño y aspectos reproductivos de E. pycnophylla en el Volcán Chiles (Nariño), comparó diferentes elevaciones, encontrando que únicamente el número de capítulos por sinflorescencia en individuos adultos varia de forma lineal negativa con la elevación. Sin embargo, la relación no fue igual para el resto de edades y el total poblacional, por lo que un juicio de generalización era muy limitado. El autor explicó que el efecto del fuego puede obscurecer las relaciones lineales propuestas por Smith (1980).
Sánchez (2004) realizó un estudio en el páramo el Ángel en Ecuador, dirigido directamente a probar la hipótesis de Smith (1980) sobre la excepción de E. pycnophylla del patrón general y a buscar la supuesta relación lineal negativa entre elevación y altura corporal. Incluyó además relaciones con otras variables morfométricas como el diámetro de la roseta y la producción reproductiva, encontrando que la única variable correlacionada linealmente con la elevación es el número promedio de capítulos por sinflorescencia, el cual disminuye con la elevación, induciendo una posible atenuación del potencial reproductivo a mayores elevaciones. Estos resultados fueron un tanto similares a los encontrados por Kovář (2001), aunque Sánchez (2004) propuso la necesidad de repetir el estudio en otros páramos de Nariño (Colombia), para buscar tendencias similares y generalizaciones. En cuanto a los patrones de supervivencia y mortalidad, Sánchez (2001) no encontró una influencia significativa por parte de la elevación, concluyendo una alta estabilidad demográfica de la especie.
Hasta el momento, estos estudios han servido para esclarecer dos asuntos relacionados con las hipótesis de Smith (1980) y proponer nuevas explicaciones alternas: 1) es cierto que E. pycnophylla es una excepción al patrón del género Espeletia, en el cual las plantas aumentan de altura corporal con la elevación; 2) no es cierta la propuesta de que en E. pycnophylla la relación sea inversa. Las investigaciones han mostrado la debilidad de tal generalización, la incompatibilidad de la linealidad con la complejidad ecológica y que las variaciones dependen de las condiciones particulares e iniciales de cada páramo, tales como la historia biológica, el efecto de la influencia antrópica y el aislamiento espacial, genético y ecológico entre poblaciones.
En este trabajo se pusieron a prueba las hipótesis de Smith (1980, 1981) y Sánchez (2004), mediante el estudio de ciertos aspectos poblacionales de E. pycnophylla ssp. angelensis en un páramo seco del departamento de Nariño y su respuesta frente al factor abiótico del gradiente altitudinal como un componente de su nicho real que regula su estructura y dinámica. Se midieron los influjos de este factor sobre un conjunto de variables y se buscaron óptimos de respuesta ecológica y explicaciones alternas en lo posible no lineales, que revelaran la importancia de otros factores sinérgicos implicados en la estructuración de la población.
Con respecto a la morfometría de adultos, el aporte de este estudio radica en que es el primero en su clase que adopta una visión multivariante para su abordaje, por lo que se puede explicar no solo la longevidad (altura de la planta) de los individuos, sino también indirectamente la importancia de dos procesos vegetales como la capacidad fotosintética, relacionada con el área foliar –dimensión de las hojas o volumen de la roseta– y la fijación de biomasa por una planta, representada en todas las variables morfométricas como su dimensión corporal.
MATERIALES Y MÉTODOS
]]> Especie de estudio. Espeletia pycnophylla es una roseta caulescente, perenne, de hojas densamente pubescentes, obovadas, oblongas, sésiles e inflorescencias desnudas. Es el único taxón dentro del género que ha sido reportado para los páramos zonales del sur de la cordillera occidental en Colombia (Nudo de los Pastos) y el Ecuador, considerándose como el último evento de especiación de la subtribu Espeletiinae. Existen tres subespecies: E. pycnophylla ssp. pycnophylla, E. pycnophylla ssp. llanganatensis y E. pycnophylla ssp. angelensis (Cuatrecasas, 1986), siendo esta última, el objeto de la presente investigación.Área de estudio. Se ubicó en la zona denominada Páramo El Infiernillo, de la Reserva Natural Pueblo Viejo, ubicada en el municipio de Mallama, departamento de Nariño (01º 03' 21' N y 77º 46' 93' O), en la zona occidental del Nudo de los Pastos. (Armero, 2004). Presenta un régimen de precipitación bimodal, con una media anual de 966 mm, un máximo de 1.361 y un mínimo de 725,6 mm. Las épocas más lluviosas son abril–mayo y octubre–noviembre, mientras que las más secas, junio, julio y agosto. La temperatura media de la zona oscila entre 10,2 y 11,3 ºC (IDEAM, 2003). El sitio se caracteriza por su alta influencia volcánica (volcanes Chiles, Cumbal y Azufral), sus suelos relativamente secos y presentar un patrón de vegetación paramuna tipo frailejonal–pajonal en su mayor extensión, dominada por las especies Espeletia pycnophylla ssp. angelensis y Calamagrostis effusa. Este páramo ha sido objeto de frecuentes quemas, la última de las cuales ocurrió en agosto del 2003, cubriendo casi completamente toda su extensión (Armero, 2004). Todo el trabajo de campo fue realizado durante la época más seca, entre julio y agosto del 2006.
Diseño y muestreo. Se cubrió un gradiente altitudinal de 600 m, desde el subpáramo hasta el páramo sensu stricto –el punto de mayor elevación del área– marcando cuatro cotas de estudio cada 200 m de elevación (3.180, 3.300, 3.500 y 3.700 m) en línea recta a modelo de transecto. Se utilizaron tres réplicas de este gradiente (T1, T2 y T3), separadas entre sí por 500 m en promedio y con las mismas características. En cada elevación se estableció una parcela de 400 m2 donde se muestrearon 30 cuadrantes de 4 m2 escogidos aleatoriamente según la metodología de Krebs (1995) y Bonilla (2003), excepto en T2 donde se muestrearon solamente nueve cuadrantes en la cota de 3.700 m debido a complicaciones de terreno. En total se evaluaron 339 cuadrantes distribuidos en toda el área de estudio. Todas las parcelas se ubicaron a favor de la pendiente en diferentes orientaciones cardinales (N, S, E, O, SO, SE, NE y NO) según el diseño aleatorio.
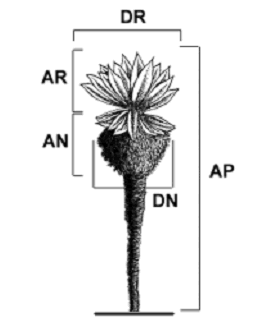
Medición de variables. Las variables medidas en cada cuadrante fueron las siguientes: 1) densidad poblacional, estimada como el número de individuos por área (# ind/4 m2); 2) Variables fenológicas como el número de sinflorescencias por individuo (NSI) y el número de capítulos por individuo (NCI), que representaron la producción de estructuras reproductivas en los adultos; 3) variables morfométricas de adultos (figura 1) como altura total de la planta (AP), altura y diámetro de su roseta (AR–DR) y altura y diámetro de su necromasa (AN–DN) (Hofstede,1995; Kovář, 2001; Sánchez, 2004), las cuales se midieron únicamente en los 25 individuos más altos y longevos por elevación; 4) mortalidad, contando el número de individuos muertos por cuadrante y realizando una estimación de su AP.
Figura 1. Variables morfométricas tomadas en individuos adultos de E. pycnophylla ssp. angelensis en el páramo el Infiernillo (AP = altura de la planta; AR = altura de la roseta; DR = diámetro de la roseta; AN = altura de la necromasa; DN = diámetro de la necromasa)
Análisis de datos. Todos los individuos –vivos y muertos– fueron clasificados en nueve clases de edad bajo dos criterios: 1) estado fenológico fértil o evidencias de eventos reproductivos pasados como sinflorescencias o sus pedúnculos secos alrededor y/o entre la necromasa –en caso de ausencia de ambas condiciones, los individuos fueron catalogados inequívocamente como juveniles– y 2) la variable AP, en intervalos regulares de 30 cm, de forma similar a lo propuesto por Armero (2004), Kovář (2001) y Sánchez (2004) para la misma especie, ya que se la ha trabajado como un estimador del tiempo de vida de estas plantas. Lo anterior se realizó con el objetivo de determinar la estructura demográfica de la población y construir curvas de supervivencia utilizando los datos derivados de una tabla de vida estática, mediante la proporción de plantas vivas y muertas en cada clase de edad (Bonilla, 2004; Sánchez, 2004). El modelo gráfico se testeó con una prueba de bondad de ajuste de Chi–cuadrado (c2) y las diferencias en las tendencias de las curvas de supervivencia entre elevaciones –representadas en sus pendientes– mediante un ANOVA (Análisis de varianza simple, F) (Zar, 1996).
El patrón de distribución espacial fue calculado con base en el índice de Morisita (Id) (Brower et al., 1998), ya que éste no es influenciado por la densidad poblacional y/o el tamaño de la unidad muestral. Las diferencias de Id entre elevaciones fueron susceptibles de ser probadas mediante un ANOVA simple más comparaciones múltiples de Tukey, gracias a que se contó con la presencia de las réplicas. La densidad poblacional y la producción de estructuras reproductivas entre elevaciones, fueron comparadas con la prueba no paramétrica de Kruskall–Wallis (H), mas comparaciones post–hoc múltiples tipo–Tukey (Zar, 1996). La relación de cada variable con el gradiente altitudinal, se probó con el índice de correlación no paramétrico de Spearman (rs). La comparación de la morfometría de individuos adultos entre elevaciones, se efectuó con los modelos multivariantes MANOVA (Análisis de varianza multivariante), pruebas post–hoc múltiples de Hotelling y un análisis de variables canónicas o discriminante (CVA), el último de los cuales se usó para identificar las variables morfométricas más importantes en caso de existir diferencias significativas. Este CVA fue realizado mediante el método de Fisher (Timm, 2002), con el que se obtuvieron funciones discriminantes canónicas que representaron la mayor variación posible con base en combinaciones lineales de las variables originales, disminuyendo la dimensionalidad de los datos y permitiendo una visualización gráfica de la variación entre elevaciones sobre un espacio bidimensional. El valor lambda de Wilks (L) fue usado como una medida del poder discriminante de las funciones (siendo 0 y 1 el máximo y mínimo poderes discriminantes respectivamente) (Álvarez, 1995).
]]> Todos los análisis estadísticos fueron realizados con el paquete Past (Paleonthological Statistics 1,38), con un nivel de significancia del 95%. Se tuvo en cuenta los requisitos de normalidad y homocedasticidad univariante –pruebas de Shapiro–Wilk y Barttlet respectivamente–, y multivariante –pruebas de Mardia y Box' M, respectivamente. Los datos morfométricos fueron transformados a logarítmos naturales cuando no cumplieron los requisitos paramétricos.
RESULTADOS
Se encontró un total de 1.156 individuos, para un área muestreada de 1.440 m2. De estos, T3 presentó la mayor abundancia con 703, seguido por T1 con 391 y T2 con 224. Del total de las plantas, 1.072 (92,73%) fueron individuos vivos y 84 (7,26%), individuos muertos. Por otra parte, 765 (66,17%) no presentaron ninguna evidencia de eventos de floración, por lo que fueron considerados como juveniles y 391 (33,82%), presentaron al menos una evidencia reproductiva y se consideraron como adultos. Ningún individuo con AP menor a 30 cm presentó evidencias de floración y todos los individuos con AP superior a 70 cm, presentaron al menos una evidencia.
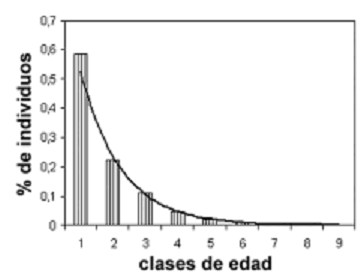
Estructura demográfica de la población de E. pycnophylla ssp. angelensis. Todos los resultados obtenidos acerca del influjo de la elevación sobre los aspectos poblacionales de E. pycnopnylla ssp. angelensis concernientes a este estudio, deben ser juzgados e interpretados con base en la estructura demográfica de la población, la cual se ajustó muy bien al modelo de la serie logarítmica (c2 = 4,71, p = 0,85). La figura 2 muestra la distribución de los individuos en las nueve clases de edad –bajo el criterio de AP–, y su aporte proporcional al total de la abundancia poblacional. La primera clase aporta el 58% de la abundancia total, y ésta desciende logarítmicamente hasta la novena clase, donde se encuentran los individuos más longevos, que aportan únicamente un 0,09%. Así, algo más de la mitad de la población está compuesta por individuos estrictamente juveniles, lo que se constituye en evidencia de un estado de regeneración temprano y una población en crecimiento.
Figura 2. Modelo demográfico logarítmico de la población de E. pycnophylla ssp. angelensis en el páramo El Infiernillo (Nariño), Colombia cuyos parámetros fueron x = 0,99 y a = 1,33
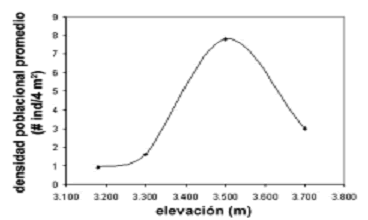
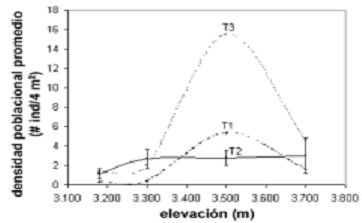
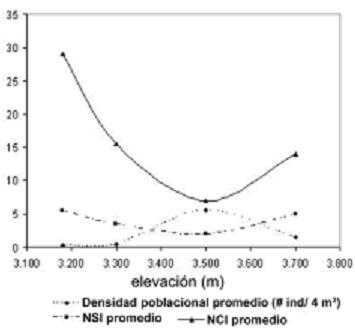
Densidad poblacional. Esta variable difirió de forma muy altamente significativa entre las cuatro elevaciones (H = 120, p < 0,001), aunque dichas diferencias fueron más leves entre 3.180 y 3.300 m (p = 0,03). Sin embargo, la variación no obedeció a relaciones lineales de ningún tipo (rs = 0,8, p = 0,2) (figura 3). Es evidente que el efecto del gradiente altitudinal sobre la densidad poblacional de E. pycnophylla ssp. angelensis describe una curva coenoclínica quasi–simétrica, con un óptimo en 3.500 m (7,8 ind/4 m2), un punto intermedio del gradiente. El mismo patrón fue observado tomando cada transecto independiente, lo que indicó una posible tendencia inequívoca para ésta zona (figura 4). En la figura 4 también puede observarse que existen diferencias entre transectos, es decir, que a parte del gradiente altitudinal, la heterogeneidad horizontal en éste páramo es un factor determinante sobre la variación de la densidad poblacional de E. pycnophylla ssp. angelensis. Las medianas de densidad fueron mayores para T3 con 5,84 ind/4 m2, seguido por T2 con 2,26 ind/4 m2 y finalmente T1 con 1,92 ind/4 m2, difiriendo de forma muy altamente significativa (H = 36,25; p < 0,001).
]]> Figura 3. Variación coenoclínica de la densidad poblacional promedio de E. pycnophylla ssp. angelensis, tomando las elevaciones comunes de las tres réplicas del gradiente altitudinal estudiado en el páramo El Infiernillo (Nariño), Colombia.
Figura 4. Variación de la densidad poblacional promedio de E. pycnophylla ssp. angelensis por cada una de las tres réplicas en el gradiente altitudinal estudiado en el páramo El Infiernillo (Nariño), Colombia. Las barras verticales en T2 indican errores estándar al 95% en cada elevación. El amplio error estándar de la cota de 3.700 m se debió a la reducción del número de cuadrantes muestreados e indica que el valor medio real podría ubicarse más abajo del observado, de acuerdo a la tendencia de las otras dos réplicas
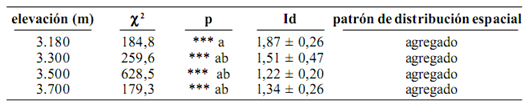
Patrón de distribución espacial. Se encontró en todo el gradiente altitudinal, que los individuos de E. pycnophylla ssp. angelensis se distribuyen de forma agregada en el espacio que ocupan, sin embargo, el grado de agregación fue significativamente más alto en 3.180 m (F = 13,96, p = 0,004) con respecto a las demás cotas, las cuales presentaron índices muy similares entre sí. Esto significa que en este punto altitudinal subparamuno, cerca al límite con el bosque, los individuos tienden a agruparse con mayor intensidad, o a depender de un mayor grado de reclutamiento. La figura 5 muestra la naturaleza de esta variación frente a la elevación, la cual posee la misma tendencia en los tres transectos.
La tabla 1 muestra los valores de la prueba de bondad de ajuste con c2 a la distribución de Poisson, la agrupación de elevaciones según la prueba a posteriori de Tukey y los valores Id, que para el caso de distribuciones agregadas, toman un rango entre 1 y el número máximo de cuadrantes muestreados.
Tabla 1. Valores de la prueba de bondad de ajuste a la distribución de Poisson. Valores medios del índice de Morisita (Id) (promedio ± error estándar 95%) y tipo de distribución de E. pycnophylla ssp. angelensis en cada elevación estudiada en el páramo El Infiernillo (Nariño), Colombia (valores de p seguidos de la misma letra, no indican diferencias significativas. Tukey 95%. Nivel de significancia *** = p < 0,001: diferencias muy altamente significativas de un patrón aleatorio)
]]>
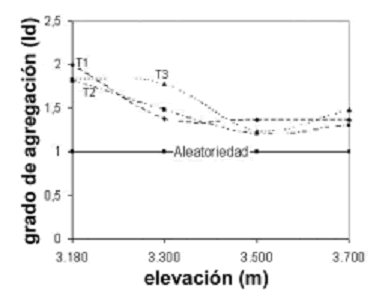
Figura 5. Variación del grado de agregación (Id) de los individuos de E. pycnophylla ssp. angelensis en el gradiente altitudinal estudiado en el páramo El Infiernillo (Nariño), Colombia. La línea continua indica el valor crítico de distribuciones aleatorias. T1 = 90 cuadrantes muestreados, T2 = 69 cuadrantes muestreados y T3 = 90 cuadrantes muestreados
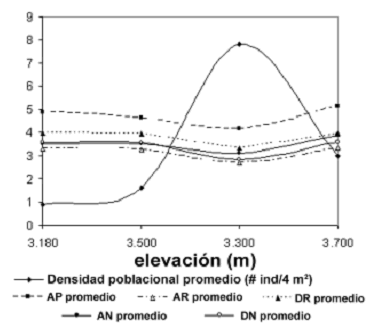
Producción de estructuras reproductivas. Entre los pares de elevaciones de 3.180–3.500 m y 3.180–3.700 m existieron diferencias muy altamente significativas en cuanto a la producción media de NCI (H = 18,68, p = 0,0003). Para NSI las diferencias se presentaron de forma significativa entre 3.180 y 3.500 m (H = 9,44, p = 0,02). A pesar de estas discrepancias, no se muestra una tendencia lineal de ambas variables frente a la elevación (NCI rs = –0,8, p = 0,2) y (NSI rs = –0,4, p = 0,6). La producción media de NCI y NSI por individuo disminuyen progresivamente desde 3.180 m y alcanzan un punto mínimo en 3.500 m, a partir del cual los valores vuelven a incrementarse hacía 3.700 m. De forma interesante, las formas de estas curvas (figura 6) son inversas a la coenoclina de densidad poblacional de la figura 4, con la misma condición quasi–simétrica. Esto empieza a mostrar explicaciones alternas acerca de que el hacinamiento y la capacidad de carga dentro de la competencia intraespecífica, son factores determinantes sobre la producción media de estructuras reproductivas en los individuos adultos de la población.
Figura 6. Variación del valor promedio de NSI y NCI en plantas adultas de E. pycnophylla ssp. angelensis en el gradiente altitudinal estudiado en el páramo El Infiernillo (Nariño), Colombia. Las barras verticales indican errores estándar al 95%
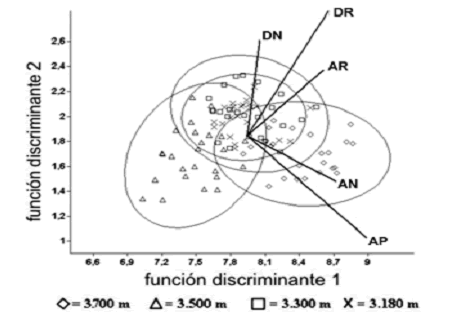
Estructura morfométrica de adultos. El MANOVA y las comparaciones múltiples post–hoc de Hotelling, permitieron inferir que la morfometría de los individuos adultos difiere de forma muy altamente significativa entre todas las elevaciones (Wilk's Lambda = 0,1737, p < 0,001), excepto entre 3.180 y 3.300 m (p = 0,85). El CVA redujo las variables originales a dos funciones discriminantes, de las cuales la primera explicó el 69,81% y la segunda el 30,01% de la variación original total, expresando en conjunto un 99,82% de la misma, la cual se consideró satisfactoria. Los eigenvectores (pesos de las variables originales dentro de la separación morfométrica de adultos entre elevaciones) (tabla 2) obtenidos con el CVA, mostraron conjuntamente que para ambas funciones discriminantes, AP fue la variable más afectada. Sin embargo, todas las dimensiones morfométricas fueron influenciadas significativamente por la elevación, por lo que se concluyó que se esta explicando diferencias en cuanto al tamaño y la forma corporal de las plantas. La tabla 3 muestra los promedios de cada variable por elevación y la figura 7 el diagrama de puntos del CVA, con la ubicación morfométrica de los 25 adultos por cota, respecto a las funciones discriminantes.
]]>

Tabla 2. Eigenvectores del CVA dentro de las dos primeras funciones discriminantes para las variables morfométricas de los individuos adultos de E. pycnophylla ssp. angelensis estudiados en el páramo El Infiernillo (Nariño), Colombia (AP = altura de la planta; AR = altura de la roseta; DR = diámetro de la roseta; AN = altura de la necromasa; DN = diámetro de la necromasa)
Tabla 3. Promedios ± error estándar (95%) de las variables morfométricas (cm) de los individuos adultos de E. pycnophylla ssp. angelensis para cada elevación en el páramo El Infiernillo, (Nariño), Colombia (AP = altura de la planta; AR = altura de la roseta; DR = diámetro de la roseta; AN = altura de la necromasa; DN = diámetro de la necromasa)
Figura 7. Diagrama de puntos del CVA. Donde muestran gráficamente la primera y segunda funciones discriminantes (ejes X e Y respectivamente) y la posición de los individuos de E. pycnophylla ssp. angelensis de cada elevación como proyecciones ortogonales de estos. Las líneas proyectadas desde el centro indican gráficamente los eigenvectores o pesos de cada variable original en cada eje o función discriminante y las elipses los intervalos de confianza del 95%. Los datos están transformados a logaritmos naturales
De los anteriores análisis, se revelaron los siguientes patrones: 1) evidentemente ninguna variable morfométrica se correlacionó de forma lineal con el gradiente altitudinal, lo que permitió rechazar inmediatamente las hipótesis generalizadas por otros autores; 2) los adultos de la elevación de 3.500 m fueron los más pequeños en todos los aspectos morfométricos, –tamaño y forma corporal de la planta– presentando las puntuaciones más bajas para todas las variables y ubicándose hacía la parte izquierda de la primera función discriminante (figura 7). De forma interesante, estas plantas están ubicadas en la cota de mayor densidad poblacional, y el gradiente en general muestra nuevamente un patrón coenoclínico de variación; 3) los adultos de 3.700 m presentaron las mayores dimensiones morfométricas, y fue ésta la única elevación donde se encontraron individuos en las clases de edad octava y novena; 4) Los adultos de las elevaciones 3.180 y 3.300 m se solaparon casi completamente en sus dimensiones morfométricas, coincidiendo además con el hecho de que sus densidades poblacionales fueron las mas símiles de todo el gradiente altitudinal. Estos resultados eliminan cualquier hipotética linealidad, pero ponen al descubierto que la variación no es totalmente errática y que exhibe la misma relación inversa con la densidad poblacional encontrada para la producción reproductiva (figura 8).
]]>Figura 8. Relación inversa entre los promedios de densidad poblacional y los promedios de NCI y NSI de E. pycnophylla ssp. angelensis en el gradiente altitudinal estudiado en el páramo El Infiernillo (Nariño), Colombia.
Lo anterior puede ser interpretado de las siguientes dos formas: a) en las zonas de elevada densidad poblacional –en este caso 3.500 m–, la cantidad de biomasa media fijada por los adultos es mínima, ya que presentan tamaños inequívocamente menores. Consiguientemente, poseen rosetas más pequeñas, lo que de forma indirecta indicaría una menor tasa fotosintética y un potencial de crecimiento limitado, sugiriendo a la competencia intraespecífica por recursos como el posible proceso causal de este patrón. La relación de la densidad poblacional con el tamaño de las plantas se muestra claramente en la figura 9; b) la probabilidad de que un individuo juvenil alcance edades avanzadas –clases de edad séptima octava y novena– en la elevación de 3.500 m es muy reducida, lo que debería verse reflejado en las tablas de vida como bajas probabilidades de supervivencia y altas probabilidades de mortalidad en las primeras clases de edad. En este caso, los efectos de la densodependencia y la competencia intraespecífica se están manifestando sobre la mortalidad de las plantas más longevas.
Figura 9. Relación inversa entre los promedios de densidad poblacional y los promedios de cada una de las variables morfométricas en adultos de E. pycnophylla ssp. angelensis en el gradiente altitudinal estudiado en el páramo El Infiernillo (Nariño), Colombia. Los datos morfométricos están transformados a logaritmos naturales con el propósito de facilitar la observación
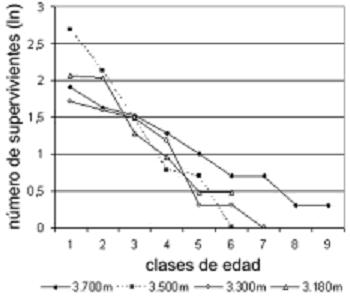
Curvas de supervivencia. Como lo muestra la figura 10, las curvas de supervivencia para todas las ]]>
elevaciones poseen la misma tendencia, un modelo tipo II (Brower et al., 1998), que explica que la probabilidad de supervivencia de los individuos disminuye proporcionalmente con su edad. Además, esto apoya el modelo de la estructura demográfica obtenida inicialmente. Las pendientes de las curvas no mostraron diferencias significativas (F = 0,85, p = 0,48), sin embargo, se pudo observar que en 3.700 m las probabilidades de supervivencia de los adultos son mayores que para el resto de cotas, permitiendo una mayor proporción de plantas longevas y contrastando sobre todo con la elevación de 3.500 m, la cual posee una curva de pendiente muy abrupta, en la que los individuos solo alcanzan la sexta clase de edad con una probabilidad de supervivencia casi nula.
Figura 10. Curvas de supervivencia de la población de E. pycnophylla ssp. angelensis para cada elevación estudiada en el páramo El Infiernillo (Nariño), Colombia. Los valores del número de supervivientes están transformados a logaritmos naturales
DISCUSIÓN
Modelo de estructura demográfica de E. pycnophylla ssp. angelensis en el páramo El Infiernillo (Nariño), Colombia. Toda discusión acerca de los efectos de la elevación sobre los aspectos autoecológicos de E. pycnophylla ssp. angelensis concernientes a esta investigación, deben basarse en el modelo demográfico logarítmico en el sito de estudio. Este indica que la población se encuentra en un temprano estado de regeneración, como respuesta al último evento de quema ocurrido hace tres años y medio. Este tipo de curvas han sido encontradas para la misma especie en Ecuador (Sánchez, 2004) y para otras especies como Espeletia hartwegiana Cuatrec. (Asteraceae) en el páramo de los Nevados (Verweij y Kok, 1992). Con base en esto, se puede corroborar la alta eficiencia y velocidad con las que este tipo de plantas se regeneran después de un evento ecológicamente desfavorable, tal como lo plantea Hofstede (2001), de manera que se podría esperar que en ausencia de un nuevo evento de fuego, la población permanecería demográficamente estable con una consecuente y mínima reducción de la abundancia de la primera clase de edad.
Efecto del gradiente altitudinal sobre la densidad poblacional de E. pycnophylla ssp. angelensis. En este caso se generó la primera evidencia empírica de la hipótesis de Smith (1980), acerca de que E. pycnophylla ssp. angelensis en los páramos colombo–ecuatorianos es una excepción a las tendencias lineales encontradas para la mayoría del género. Aquí no se encontró una relación lineal entre la elevación y la densidad, sin embargo, la naturaleza de la variación no es totalmente indiferente al gradiente o errática. Lo que se evidenció es un fenómeno muy interesante y estudiado en la ecología de poblaciones: una variación coenoclínica. Existe un óptimo de tolerancia altitudinal (nivel intermedio del factor), donde la abundancia de los individuos es máxima, pero disminuye quasi–simétricamente hacia ambos lados del gradiente. Una de las posibles razones de lo anterior puede ser la heterogeneidad topográfica, ya que la elevación de 3.500 m es una zona de pendientes relativamente leves con respecto a las demás. Smith (1981) menciona que los suelos en este tipo de zonas presentan una mayor acumulación de agua, nutrientes y menor escorrentía, lo que favorece una densidad poblacional alta. Esto se debe a que la probabilidad de que una semilla germine en estas condiciones es mayor que en zonas con pendientes más fuertes y menor disponibilidad hídrica. Sánchez (2004) también rechaza la hipótesis de linealidad en cuanto a la naturaleza de la variación de la densidad poblacional frente a la elevación, aunque no obtuvo la misma respuesta encontrada en este trabajo. Además, hay que tener en cuenta la historia natural y de disturbio de cada zona de páramo, la cual varía inclusive en escalas espaciales muy pequeñas, lo que hace imposible establecer juicios de generalización verosímiles para una especie en todo su ámbito geográfico.
Efecto del gradiente altitudinal sobre el patrón de distribución espacial. Los individuos de E. pycnophylla ssp. angelensis tienden a agregarse más intensamente a menores elevaciones, donde la densidad poblacional es baja, mientras que en áreas muy densas la agregación es más leve. Esto puede ser explicado desde dos puntos de vista: 1) simplemente se trata de un artificio estadístico y de muestreo, ya que debido al fenómeno del reclutamiento en esta especie, la probabilidad de encontrar cuadrantes vacíos es obviamente muy alta en áreas poco densas, lo que generalmente ocurre en la transición ecotónica bosque alto andino–subpáramo; 2) el reclutamiento en esta especie es considerado como una estrategia para que la descendencia de un individuo tenga una alta probabilidad de germinar en zonas contiguas a su progenitor. Desde el punto de vista de la competencia interespecífica, en zonas de alta densidad poblacional, donde la especie es relativamente dominante, no se necesitan altas tasas de reclutamiento, ya que en cualquier punto espacio–temporal, las semillas tienen una alta posibilidad de germinar. En zonas de baja densidad poblacional, donde no existe dominancia notable, las plantas se ven presionadas por la competencia de otras especies con requerimientos similares o solapados en cualquier dimensión de su nicho hiper–espacial, por lo que la estrategia ecológica para asegurar que la energía gastada en producción reproductiva no se pierda, es incrementar el grado de reclutamiento. Así, las semillas que tienen mayor probabilidad de germinar son las que caen en las zonas circundantes a sus progenitores.
Efecto del gradiente altitudinal sobre las variables fenológicas NSI y NCI. Sánchez (2004) encontró una relación supuestamente inequívoca, en la que la producción reproductiva media por individuo disminuye con la elevación, y generalizó la relación para otras zonas de distribución de la especie, incluso para el departamento de Nariño. Fue interesante encontrar que en este estudio la variación de NSI y NCI frente a la elevación se comporta como una 'coenoclina invertida', en la que existe un punto crítico de producción reproductiva en niveles intermedios del factor y un aumento quasi–simétrico hacia ambos lados del gradiente altitudinal. Esta relación es exactamente inversa a la variación de la densidad poblacional, o sea que en zonas de mayor abundancia de individuos por área, hay una mayor efectividad reproductiva con menor inversión en la producción de estructuras por individuo. ¿Es esto una paradoja ecológica? Se debería esperar lo contrario, que una de las posibles explicaciones de la alta densidad poblacional en un espacio determinado sea que los individuos allí presentes, produzcan una alta cantidad de estructuras reproductivas como flores, capítulos y sinflorescencias. Sin embargo, se necesitan otros estudios que traten de dar respuesta a esta cuestión aparentemente contradictoria.
]]> Efecto del gradiente altitudinal sobre la morfometría de adultos. Este componente arroja la segunda evidencia empírica de que la hipótesis de Smith (1980) sobre relación lineal negativa entre el tamaño de las plantas adultas de E. pycnophylla ssp. angelensis y la elevación, no puede ser generalizada. Otros autores como Kovář (2004) y Sánchez (2004), tampoco encontraron en la misma especie una relación clara entre la altura de los individuos adultos y la elevación. Nuevamente se elucida un interesante patrón que tiene una relación inversa con la densidad poblacional. Los individuos que se encuentran en zonas de elevada densidad poblacional son inequívocamente más pequeños en todas sus dimensiones morfométricas. Esto puede ser explicado por medio de dos perspectivas: 1) los adultos están siendo limitados en su crecimiento debido a la alta competencia intraespecífica por espacio y nutrientes que sufren en una zona muy densa. Por lo tanto, la asimilación de biomasa (representada indirectamente en tamaño total de la planta) es una variable densodependiente; 2) observando que en 3.500 m la mayoría de los adultos están ubicados alrededor de las clases de edad cuarta (91–120 cm) y quinta (121–150 cm), la probabilidad de que un individuo alcance una elevada longevidad (la máxima para esa población), y por lo tanto una talla corporal grande, es muy reducida. Nuevamente aquí, la explicación es la densodependencia y la competencia intraespecífica. Por lo tanto no se puede generalizar la hipótesis de Smith (1980). Más bien, esta relación está determinada por muchos otros factores que interactúan de forma compleja, dentro de los cuales podemos mencionar a la competencia intraespecífica aquí encontrada y a los daños impredecibles que en el tiempo causa la historia de disturbio de cada páramo.La curva de supervivencia de la población de E. pycnophylla ssp. angelensis y su variación frente al gradiente altitudinal; lo que se puede inferir de este componente, es que debido al temprano estado de regeneración de la población de E. pycnophylla ssp. angelensis en el páramo el Infiernillo (Nariño), Colombia, la elevación no influye de manera significativa sobre los patrones de mortalidad y supervivencia de los individuos de diferente edad. Las tendencias siempre son similares ya que hay que considerar que estamos hablando de un spatium–contiinum de una población completa en donde los criterios de separación que proponen los investigadores no dejan de tener cierta arbitrariedad. En general, los patrones de mortalidad y supervivencia de esta población no se ven influenciados por la elevación.
AGRADECIMIENTOS
Los autores agradecemos la financiación completa de este trabajo al sistema de investigaciones de la Universidad de Nariño, al Biól. Msc. Belisario Cepeda por su colaboración en todas las fases de la investigación, al grupo de investigación Biología de Páramos y Ecosistemas Andinos de la Universidad de Nariño, a la Reserva Natural Pueblo Viejo por permitirnos la realización del trabajo de campo y a los evaluadores del manuscrito.
REFERENCIAS
1. Álvarez F. 1995. Estadística multivariante con SPSS. Aplicación a las ciencias de la salud. Ediciones Díaz de Santos, S.A. Madrid, España. [ Links ]
2. Armero SM. 2004. Efecto de la quema sobre algunos aspectos poblacionales de Espeletia pycnophylla Cuatrec. en el páramo El Infiernillo (Mallama–Nariño). Tesis de pregrado. Programa de Biología, Facultad de Ciencias Naturales y Matemáticas, Universidad de Nariño. San Juan de Pasto (Nariño), Colombia. [ Links ]
3. Bonilla MA. 2004. Ecología de poblaciones. Universidad Nacional de Colombia, Facultad de Ciencias, Departamento de Biología. Bogotá D. C., Colombia. [ Links ]
4. Bonilla MA, Guillot G. 2003. Prácticas de ecología. Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia. Bogotá D. C., Colombia. [ Links ]
5. Brower JE, Zar JH, Von Ende CN. 1998. Field and laboratory methods for general ecology. Fourth Edition. McGraw Hill. Boston, U. S. A. [ Links ]
6. Cuatrecasas J. 1986. Speciation and radiation of the Espeletiinae in the Andes. Pp. 267–303. En: Vuilleumier F, Monasterio M (eds.). High altitude tropical biogeography. Oxford University Press. New York, U. S. A. [ Links ]
7. Hofstede RG. 1995. Effects of burning and grazing on a Colombian paramo ecosystem. PhD. Dissertation. University of Ámsterdam. 119. Ámsterdam, Netherlands. [ Links ]
8. Hofstede RG. 2001. El impacto de las actividades humanas sobre el páramo. Pp. 113–118. En: Mena V, Medina G, Hofstede RG (eds). Los páramos del Ecuador, particularidades, problemas y perspectivas. Proyecto Páramo. Quito, Ecuador. [ Links ]
9. IDEAM. 2003. Reporte de precipitación de 21 años de mediciones en la estación meteorológica El Paraíso, Ipiales, Nariño. IDEAM. Colombia. [ Links ]
10. Kovář P. 2001. Effects of burning on Espeletia pycnophylla stands in the paramo of Volcán Chiles, Ecuador –a short–term case. Pp. 113–118. En: Ramsay PM (ed.). The ecology of Volcan Chiles: high altitude ecosystems on the Ecuador–Colombia border. Pebble & Shell. Plymouth. U. K. [ Links ]
11. Krebs CJ. 1989. Ecological methodology. Harper & Row. New York, U. S. A. [ Links ]
12. Meinzer FC, Goldstein GH, Rundel PW. 1985. Morphological changes along an altitude gradient and their consequences for an Andean giant rosette plant. Oecología, 65:278–283. [ Links ]
13. Sánchez A. 2004. Análisis morfométrico y demográfico de Espeletia pycnophylla Cuatrecasas. en un gradiente altitudinal provincia de Carchi–Ecuador. Tesis de pregrado. Programa de Ciencias biológicas, Facultad de Ciencias, Universidad de los Andes. Bogotá D. C., Colombia. [ Links ]
14. Smith AP. 1980. The Paradox of plant height in an Andean giant rosette species. Journal of Ecology, 68:63–73. [ Links ]
15. Smith AP. 1981. Growth and population dynamics of Espeletia (Compositae) in the Venezuelan Andes. Smithsonian Contributions to Botany, 48:1–45. [ Links ]
16. Sokal RR, Rohlf FJ. 1995. Biometry. 3th. Edition. Freeman and Company. New York, U. S. A. [ Links ]
17. Timm, NH. 2002. Applied multivariate analysis. Springer. New York, U. S. A. [ Links ]
18. Verweij PA, Kok K. 1992. Effects of fire and grazing on Espeletia hartwegiana populations. Pp. 215–229. En: Balslev H, Luteyn L (eds.). Páramo: An andean ecosystem under human influence. Academic Press. London, U. K. [ Links ]
19. Zar HJ. 1996. Biostatistical analysis. 3th. edition. Editorial Prentice Hall. NJ, U. S. A. [ Links ]
Recibido: agosto 2007; aceptado: noviembre 2007. ]]>