CARACTERÍSTICAS BIOLÓGICAS DEL BLANQUILLO SORUBIM CUSPICAUDUS LITTMANN, BURR Y NASS, 2000 Y BAGRE RAYADO PSEUDOPLATYSTOMA MAGDALENIATUM BUITRAGO-SUÁREZ Y BURR, 2007 (SILURIFORMES: PIMELODIDAE) RELACIONADAS CON SU REPRODUCCIÓN EN LA CUENCA MEDIA DEL RÍO MAGDALENA, COLOMBIA
BIOLOGIC CHARACTERISTICS OF BLANQUILLO SORUBIM CUSPICAUDUS LITTMANN, BURR Y NASS, 2000 AND BAGRE RAYADO PSEUDOPLATYSTOMA MAGDALENIATUM BUITRAGO-SUÁREZ Y BURR, 2007 (SILURIFORMES: PIMELODIDAE) RELATED TO THEIR REPRODUCTION IN THE MIDDLE BASIN OF THE MAGDALENA RIVER, COLOMBIA
Luz F. Jiménez Segura1; Jaime Palacio2; Raúl López3
1 Docente. Instituto de Biología, Universidad de Antioquia. Medellín (Antioquia), Colombia. ljimenez@matematicas.udea.edu.co
2 Ingeniería Sanitaria, Universidad de Antioquia. Medellín (Antioquia), Colombia. japalaci@udea.edu.co
]]> 3 Departamento de Ecología, Universidad Militar Nueva Granada. Bogotá, Colombia. rlopezp@umng.edu.co
Resumen
Algunas características biológicas de Pseudoplatystoma magdaleniatum y de Sorubim cuspicaudus fueron monitoreadas durante los años 2004 y 2005 en la cuenca media del río Magdalena y, analizamos si estas se encuentran sincronizadas con la densidad de larvas de estas dos especies derivando en el río Magdalena. Para esto se utilizó la información de 1.245 individuos adultos de P. magdaleniatum y 1.206 de S. cuspicaudus y la densidad de sus larvas en muestras de agua del río Magdalena. S. cuspicaudus se reproduce durante el periodo de ''aguas bajas'' mientras que P. magdaleniatum lo hace durante el periodo de ''aguas subiendo''. Las hembras de las dos especies fueron siempre de mayor tamaño que los machos, la relación longitud-peso fue alométrica y solo en el caso de P. magdaleniatum las características biológicas estuvieron sincronizadas con la densidad de larvas durante el periodo de observación.
Palabras clave: larvas, Pimelodidae, reproducción, río Magdalena
Abstract
Some biological characteristics of Pseudoplatystoma magdaleniatum and Sorubim cuspicaudus were monitored during years 2004 and 2005 in the middle basin of the Magdalena river, and we analyzed if they were synchronized with their larvae densities drifting in the Magdalena river. We use data from 1245 adults of P. magdaleniatum and 1206 of S. cuspicaudus and larvae densities from water samples of the Magdalena river. S. cuspicaudus reproduces in the ''low water'' hydro-period and P. magdaleniatum does during ''high water'' hydro-period. Females were always bigger than males. Lenght-weigth relationship was allometric and only the biologic characteristics of adults of P. magdaleniatum were associated with their larvae densities.
Key words: Pimelodidae, reproduction, fish larvae, Magdalena river
]]>
INTRODUCCIÓN
La actividad pesquera en el río Magdalena se sustenta en poblaciones de múltiples especies y esta estrechamente asociada a los movimientos de los peces migratorios como el bocachico Prochilodus magdalenae, el bagre Pseudoplatystoma magdaleniatum, el blanquillo Sorubim cuspicaudus, la dorada Brycon moorei, la picuda Salminus affinis, entre las más importantes (Valderrama-Barco y Zarate-Villareal 1989). El bagre y el blanquillo son especies muy apreciadas en el mercado nacional por la calidad de su carne y representan el 8 y 5%, respectivamente, en las capturas de los pescadores artesanales (CCI-INCODER, 2007).
El río Magdalena es el principal sistema fluvial de Colombia y uno de los complejos acuáticos más extensos de Suramérica. La producción pesquera de la cuenca es de las más importantes del país; del total de la producción pesquera nacional en el año 2007 (104.805 ton), la captura continental representó cerca del 18% y, de este porcentaje, la cuenca del río Magdalena aportó el 53% (CCI-INCODER 2007). Entre los años 1995 y 2006, la productividad pesquera ha descendido a una tasa de 1% anual, llegando a volúmenes de captura de 6.044 ton en el 2006. Y el bocachico siempre ha sido la especie más importante en la pesquería pues contribuye con más del 50% de la captura total. Según los monitoreos realizados por el Estado colombiano, las especies que más reducción han tenido en las capturas han sido el bagre y el capaz (Pimelodus grosskopffi) pues su captura se ha reducido en un 55 y 90%, respectivamente (CCI-INCODER 2006). Ésto hace que los estudios dirigidos a evaluar el estado de sus poblaciones y a implementar acciones de manejo como definición de temporadas de veda a la pesca, tallas mínimas de captura y la protección de hábitats vitales en el desarrollo de su ciclo de vida, sean definitivos para encaminar acciones dirigidas a su protección y explotación sostenible.
La definición de periodos de veda a la pesca se basa en la identificación de meses de mayor actividad reproductiva de las poblaciones de especies explotadas por la pesquería. Para definir estos tiempos de actividad reproductiva son utilizados algunos indicadores indirectos como la relación gonadosomática, el factor de condición somático, la proporción sexual (Vazzoler 1996) e incluso, el coeficiente alométrico de la población observada (Wooton 1999); sin embargo, estas relaciones no necesariamente indican la ocurrencia de desoves, evento que puede ser claramente definido por el incremento en la concentración de ictioplancton en la masa de agua (Nakatani et al. 2000).
Jiménez-Segura (2008) encontró que el aumento en la densidad de larvas de algunas especies de peces migratorias en la cuenca media del río Magdalena está claramente asociado con las condiciones hidrológicas del sistema y que, estas condiciones cambian anualmente debido a las condiciones climáticas globales; lo que repercute directamente en el momento en que las especies desovan. Utilizando información biológica de individuos adultos de P. magdaleniatum y de S. cuspicaudus obtenida durante los años 2004 y 2005 en la cuenca media del río Magdalena, se analizó si sus características relacionadas con la reproducción se encuentran sincronizadas con la densidad de sus larvas en este sistema acuático.
MATERIALES Y MÉTODOS
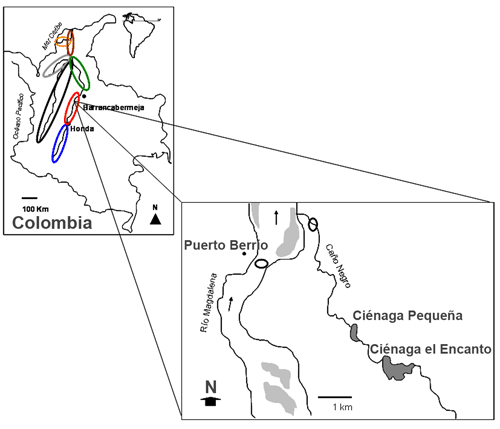
Área de estudio. La cuenca del río Magdalena, con 1.500 km de longitud, se orienta entre la latitud 3o S y 11º N. Su área de drenaje es de 257.000 km2 y descarga en el mar Caribe un volumen promedio de 6.800 m3/s. Los muestreos fueron realizados frente al municipio de Puerto Berrío (Antioquia), Colombia, sector medio de la cuenca, que se encuentra en los 111 m.s.n.m y entre los 6º 29' 15,7'' N y 74º 24' 21'' O (figura 1). Tiene una precipitación media anual de 2.200 mm. Las épocas de lluvias se observan entre abril-junio y septiembre-noviembre y los estiajes entre diciembre-marzo y entre julio y agosto. La ciudad de Puerto Berrío recoge el 24% a la captura total de la pesquería en la cuenca media, la cual aporta el 34% del volumen total de la cuenca magdalénica (INCODER-CCI 2007).
Información de adultos. La información de adultos de P. magdaleniatum y S. cuspicaudus fue registrada a partir de las capturas de pescadores de la Asociación de Pescadores de Puerto Berrío (ASOPESCA) entre agosto del 2004 y agosto del 2005. Cada individuo fue pesado (g) –peso eviscerado– y medido en su longitud estándar (cm) con precisión 0,1. El sexo y estadio de desarrollo gonadal fueron definidos mediante observación macroscópica de la cavidad visceral. Las categorías de desarrollo gonádico fueron definidas microscópicamente según la propuesta de Vazzoler (1996). Adicionalmente, se fijaron muestras de tejido de los ovarios en solución Gilson para facilitar el conteo de ovocitos.
Información de larvas. El muestreo de ictioplancton fue semanal, antes de las 8:00 horas y en dos sitios: cauce principal del río Magdalena frente a la ciudad de Puerto Berrío (Antioquia) y Caño Negro –conexión del sistema cenagoso lateral con el cauce del río Magdalena-. En el río, se seleccionaron dos ambientes de acuerdo con la margen de sedimentación: margen derecha (margen de depósito) y margen izquierda (margen de socavación). En la margen derecha la muestra fue superficial -0,5 m- y en la izquierda a dos profundidades: 0,5 m y 10 m.
Para capturar las larvas (ictioplancton) de estas dos especies se utilizó una red cónico-cilíndrica de boca circular con diámetro de 0,38 m, largo 1,2 m y de ojo de malla 0,4 mm, con un flujómetro instalado para estimar el volumen de agua filtrado. En el cauce principal del río Magdalena, las muestras se tomaron a contraflujo y, debido a la velocidad en el flujo del agua, con un tiempo de filtrado de un minuto (margen izquierda) y tres minutos (margen derecha). En Caño Negro, se realizaron arrastres en zig-zag durante cinco minutos. Cada muestra se fijó en formaldehído al 5%, neutralizada con carbonato de calcio.
Las larvas fueron separadas de material acompañante y contadas de acuerdo con su fase de desarrollo según los criterios de Kendall et al. (1984), con modificaciones de Nakatani et al. (2001) en: larval vitelino, pre-flexión, flexión y post-flexión. La identificación de las larvas fue definida a partir de diversas fuentes (Contreras y Contreras 1989, Novoa y Cataño 2005, Nakatani et al. 2001, Padilla et al. 2001).
Análisis de los datos. Ciclos y periodos hidrológicos. De acuerdo con los cambios en el promedio mensual de nivel del agua en el río Magdalena, se definieron dos ciclos hidrológicos intra-anuales (I: entre enero y junio y II: entre julio y diciembre) y cuatro periodos hidrológicos dentro de cada uno de ellos (aguas bajas, aguas creciendo, aguas altas, aguas bajando) (figura 2). La información biológica proveniente de los adultos y de las larvas fue agrupada de acuerdo con estos ciclos y periodos hidrológicos.
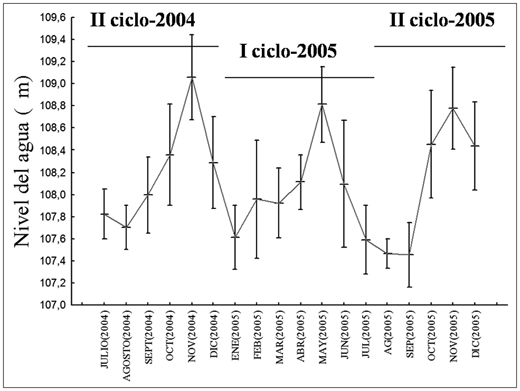
Figura 2. Distribución temporal del nivel del agua (promedio ± desviación estándar) en la cuenca media del río Magdalena durante los años 2004-2005. Circulo gris-línea continua: aguas bajas, circulo gris-línea punteada: aguas subiendo, círculo negro-línea continua: aguas altas, círculo negro-línea punteada: aguas bajando.
Información de adultos. Para cada especie en cada periodo hidrológico fueron definidos los estadígrafos de tendencia central y de dispersión de la longitud estándar y del peso total. La diferencia de estas variables entre meses y sexos fue explorada mediante un análisis de varianza. A partir de la relación longitud-peso se obtuvo la ecuación que describe el patrón de crecimiento de cada especie; para definir si éste fue diferente entre los periodos hidrológicos, se compararon las pendientes según Zar (1996). Adicionalmente, se utilizó el factor de condición alometrico k = Peso ÷ Longitud b (Ricker 1975) como expresión del bienestar de los individuos de la muestra. La proporción sexual total y mensual fue estimada de acuerdo con Vazzoler (1996). Las diferencias en las proporciones totales y mensuales, fue establecida mediante una prueba de Chi cuadrado (Cochran 1952).El tipo de desove de cada especie fue determinado a partir de la distribución de frecuencias del tamaño de los ovocitos. Para definir si existían diferencias significativas en el número de ovocitos por gramo de hembra entre los dos ciclos hidrológicos anuales, se aplico una prueba de Mann-Whitney (Man y Whitney 1947).
Información de larvas. Para permitir comparaciones entre momentos de captura, la densidad de larvas de cada especie fue estimada para un metro cúbico de agua filtrada. Se estimó la densidad de larvas en cada periodo hidrológico y ciclo intra-anual, a partir de las muestras obtenidas semanalmente durante ellos. Para definir si existieron diferencias en la densidad de larvas entre los dos ciclos hidrológicos intra-anuales se aplicó la prueba de Mann-Whitney (Man y Whitney 1947) y entre periodos hidrológicos, la prueba de Kruskal-Wallis (Kruskal 1952) según el caso.
]]> La relación entre los descriptores del estado reproductivo y de bienestar de los adultos (factor de condición k y porcentaje de individuos desovantes y su proporción sexual) y la densidad de larvas en cada periodo hidrológico fue explorada mediante análisis gráfico. El porcentaje de adultos desovantes se considera como aquella fracción de la muestra de individuos totales en cada periodo y ciclo hidrológico, que presentaron gónadas en estadio C (maduras) y D (desovadas).El momento hidrológico de ingreso de las larvas a Caño Negro, fue definido a partir del análisis gráfico de la distribución de la densidad de larvas capturadas en este ambiente en cada periodo hidrológico. La dominancia de alguna de las fases de desarrollo larval fue determinada a partir de la distribución de la densidad de cada una de ellas en cada periodo hidrológico.
A partir del tiempo que toma un individuo de estas especies de bagres para alcanzar su periodo larval (fase de flexión) desde la fertilización del ovocito y de la velocidad del agua promedio en el canal principal del río Magdalena durante cada periodo hidrológico (Jiménez-Segura 2007), se estimó la distancia aproximada que ha recorrido la prole desde la fertilización. Con base en esta distancia se define la localización teórica del área de desove. El tiempo que toma S. cuspicaudus y P. magdaleniatum para llegar a esta fase es de 62 y 110 horas post-fertilizacion, respectivamente, en rangos de temperatura entre 27 y 28,5 ºC (Contreras y Contreras 1987, Novoa y Cataño 2005, Padilla et al. 2001). Dado que la temperatura y velocidad del agua son factores físicos que pueden influir sobre la velocidad de desarrollo morfológico, se asume que estos valores se mantienen constantes durante la deriva de los individuos.
Los análisis fueron realizados en STATISTICA 6.0. Para todas la pruebas de hipótesis se asumió un nivel de significancia del 95% (p < 0,05).
RESULTADOS
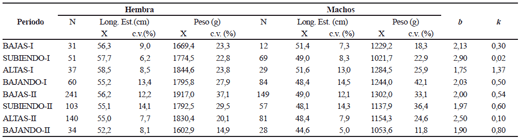
Sorubim cuspicaudus. La longitud estándar y el peso promedio de los 1206 individuos registrados durante el tiempo de estudio fue de 52,8 ± 6,9 cm y 1.562,8 ± 581,5 g, respectivamente. Las hembras presentaron mayor longitud que los machos (F (1, 1190) = 244,6; p < 0,05) y, en particular, durante las crecientes del primer ciclo anual (tabla 1). Las hembras con mayor talla se observaron en la temporada de aguas subiendo y aguas altas del primer ciclo (F(7; 1190) = 7, 46; p < 0,05).
El factor de bienestar de la población fue menor en los periodos de aguas subiendo (primer ciclo) y altas (segundo ciclo) y los mayor en las aguas altas (primer ciclo) y bajando (segundo ciclo) (tabla 1).
Tabla 1. Promedio (x) y coeficiente de variación (c. v.) de la longitud estándar (Long. est.) y peso de Sorubim cuspicaudus de acuerdo con el sexo y el periodo hidrológico durante los años 2004-2005 en el río Magdalena. N = número de individuos, k = factor de bienestar, b = coeficiente de alometría.
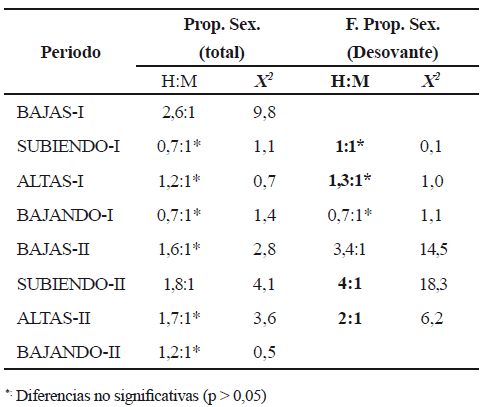
En la muestra total del número de individuos, dominaron hembras (1,3 H: 1 M) y excepto, en el periodo Bajas-I y Subiendo-II, la diferencia no fue significativa. En la fracción de adultos desovantes, no se encontraron diferencias en la proporción sexual durante el primer ciclo hidrológico pero si en el segundo ciclo, cuando el número de hembras fue muy superior al de los machos (tabla 2).
Tabla 2. Proporción sexual (Prop. Sex.) de la población total y de la fracción desovante (Estadios de madurez ovariana C y D) de Sorubim cuspicaudus en la cuenca media del río Magdalena (H = hembras; M = machos)
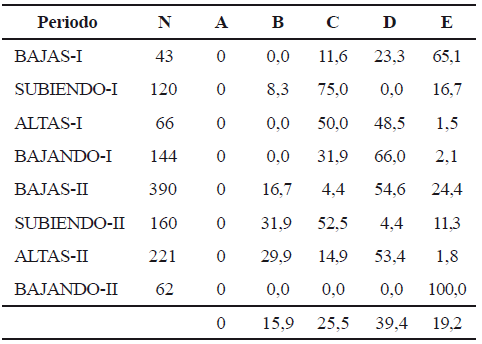
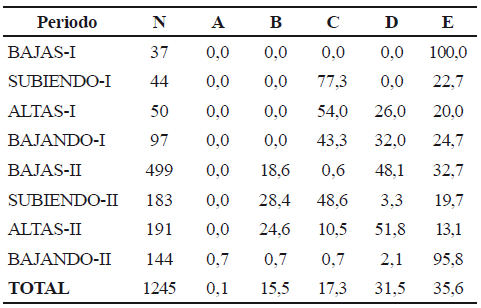
Se observaron diferentes estadios de madurez gonadal durante el tiempo de muestreo (tabla 3). Dentro de cada uno de los dos ciclos intra-anuales, el mayor porcentaje de individuos próximos a desovar (estadio C) se observó en los periodos de aguas subiendo I y II. En todos los periodos de observación, se encontraron individuos con gónadas en reposo (E).
Tabla 3. Distribución porcentual de los ejemplares de Sorubim cuspicaudus de acuerdo con la madurez de sus ovarios, durante los años 2004 y 2005 (A: virgen; B: adulto en maduración; C: adulto; D: adulto desovado; E: adulto en reposo)
Hembras con tallas entre 52 y 64 cm de longitud estándar, presentaron un promedio de ovocitos totales fue 187.959,7 (± 82.943,7) y de ovocitos maduros fue 78.943 (± 34.836). El número de ovocitos maduros por gramo de hembra fue 42,7 ovocitos g-1 (± 18,7). No se encontraron diferencias significativas en el número de ovocitos entre los dos ciclos hidrológicos intra-anuales (p = 0,49). La clase de diámetro dominante fue 0,91 a 1 mm. El diámetro promedio fue 0,89 ± 0,13 mm (n = 2.289).
Tabla 4. Densidad de larvas (individuos/m-3) de Sorubim cuspicaudus de acuerdo con el ambiente (cauce principal del río Magdalena y Caño Negro) y el periodo hidrológico durante los años 2004-2005 en el río Magdalena (X = promedio; c.v. = coeficiente de variación)
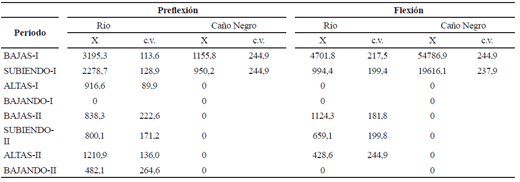
La densidad de larvas no fue diferente entre los dos ciclos intra-anuales (Z = -1,24; p = 0,21). Entre periodos hidrológicos la mayor densidad se observó durante la aguas bajas (H (7, 106) = 15,2; p = 0,03) (tabla 4). La mayor densidad de larvas en Caño Negro fue capturada solo durante el inicio del primer ciclo hidrológico intra-anual y la fase de desarrollo larval que dominó fue flexión.
De acuerdo con el tiempo de desarrollo del individuo hasta alcanzar su fase de flexión en el periodo larval y la velocidad promedio del agua, teóricamente, las zonas de desove se encuentran a una distancia teórica promedio de 131,8 km (± 9,8) aguas arriba del sitio de captura.
La relación entre proporción sexual, porcentaje de individuos desovantes y factor de condición parece no tener una relación clara con la densidad de larvas (figura 3).
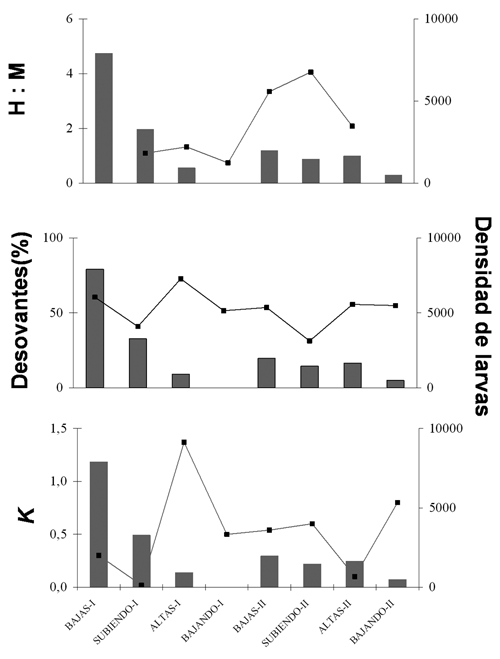
Figura 3. Porcentaje y proporción sexual de individuos desovantes y factor de condición de los adultos en relación con la densidad de larvas de Sorubim cuspicaudus en cada periodo hidrológico observado en el río Magdalena durante los años 2004-2005. Densidad de larvas en barras grises.
Pseudoplatystoma magdaleniatum. La longitud estándar promedio de los 1.245 de P. magdaleniatum n. sp. examinados fue 75,3 cm (± 17,6) y el peso promedio fue 5.060,9 g (± 3.745,7). Las hembras siempre tuvieron mayor longitud que los machos (F (1; 1229) = 475,1; p < 0,05) (tabla 5).
Tabla 5. Promedio (x) y coeficiente de variación (c. v.) de la longitud estándar (Long. est.) y peso de Pseudoplatystoma magdaleniatum de acuerdo con el sexo y el periodo hidrológico durante los años 2004-2005 en el río Magdalena (N = número de individuos; k = factor de bienestar; b = coeficiente de alometría)
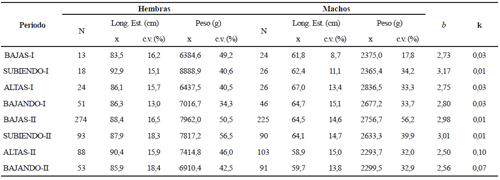
La relación longitud-peso se describe por la ecuación Peso = 0,022 * (LE) 2,82 (R2 = 0,91), cuya pendiente fue diferente entre periodos hidrológicos (t-test (1, 1.238) = 7,73; p < 0,05). Por otro lado, el factor de bienestar k presentó los menores valores en los periodos de aguas subiendo y los mayores en las altas y bajando (tabla 5).
La proporción sexual en la muestra total fue 1:1 y se conservó así durante casi todo el tiempo de observación; solo en el periodo Bajas I, los machos dominaron en número (tabla 6). Excepto durante las aguas altas, cuando los machos duplicaron en número a las hembras, en los otros periodos hidrológicos la proporción sexual no fue diferente.
Tabla 6. Proporción sexual (Prop. Sex.) de la población total y de la fracción desovante (Estadios de madurez ovarica C y D) de Pseudoplatystoma magdaleniatum en la cuenca media del río Magdalena (H = hembras; M = machos)
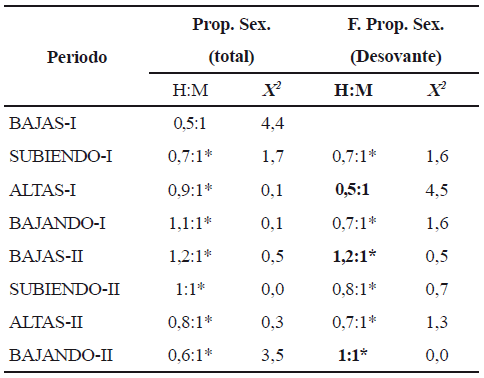
*: Diferencias no significativas (p > 0,05)
El mayor porcentaje de individuos maduros se observó durante los periodos de aguas subiendo I y II. (tabla 7). Se observaron individuos con gónadas en reposo (E) en todos los periodos hidrológicos, en particular durante los periodos bajas I y bajando II.
Tabla 7.Distribución porcentual de los ejemplares de Pseudoplatystoma magdaleniatum de acuerdo con la madurez de sus ovarios, durante los años 2004 y 2005 (A: virgen; B: adulto en maduración; C: adulto; D: adulto desovado; E: adulto en reposo)
En hembras con talla entre 82 y 115 cm, se encontró un promedio de ovocitos totales de 866232,5 (± 602052,2), el promedio de ovocitos maduros fue 493.752,5 (± 343.169,7) y por gramo fue 52,2 (± 28,8). El número de ovocitos maduros fue mayor (108,6 ovocitos g-1) en el primer ciclo hidrológico que en el segundo (p = 0,03). La clase de diámetro dominante fue de 0,91 a 1 mm. El diámetro promedio fue 0,9 mm (± 0,13) (n = 1.603).No se encontró diferencias en la densidad de larvas entre los dos ciclos intra-anuales (Z = -0,53; p = 0,59) ni entre periodos hidrológicos (H (7, 106) = 12,5; p = 0,08). A pesar de ésto, se observó la mayor densidad en el periodo de aguas bajas del primer ciclo intra-anual (tabla 8). La mayor densidad de larvas en Caño Negro se encontró en las aguas altas del primer ciclo intra-anual y en fase de flexión.
]]> Tabla 8. Densidad de larvas (individuos/m-3) de Pseudoplatystoma magdaleniatum de acuerdo con el ambiente (cauce principal del río Magdalena y Caño Negro) y el periodo hidrológico durante los años 2004-2005 en el río Magdalena (X = promedio; c.v. = coeficiente de variación)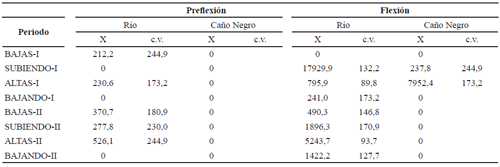
De acuerdo con el tiempo que toma un individuo hasta alcanzar su fase de flexión y con la velocidad promedio del agua, se define que, teóricamente, las áreas de desove se localizaron a 324 km (± 20,9) aguas arriba del lugar de captura.
La relación entre proporción sexual, porcentaje de individuos desovantes y factor de condición parecen estar asociados con la densidad de larvas (figura 4). La densidad de larvas se eleva cuando las proporciones sexuales 1H:1M, el incremento en porcentaje de adultos desovantes y la caída en el factor de condición. Sólo en el periodo altas-II, la relación entre el factor de bienestar no estuvo asociada con el incremento en la densidad larval.
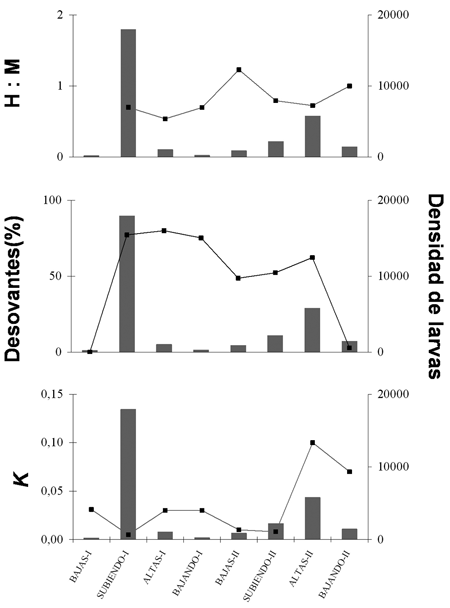
Figura 4. Porcentaje y proporción sexual de individuos desovantes y factor de condición de los adultos en relación con la densidad de larvas de Pseudoplatystoma magdaleniatum en cada periodo hidrológico observado en el río Magdalena durante los años 2004-2005. Densidad de larvas en barras grises
DISCUSIÓN
La mayoría de las tallas de captura de S. cuspicaudus y de P. magdaleniatum realizadas por los pescadores en la ciudad de Puerto Berrío se encontraron sobre la talla mínima de captura (TCM) reglamentada para la cuenca (45 cm para S. cuspicaudus y 80 cm para P. magdaleniatum); en el caso de P. magdaleniatum el 60% de lo capturado se encontró por debajo de la TMC y una amplia variación en el tamaño capturado (C. V. = 23%). Nuestros resultados hacen evidente que, contrario a la del blanquillo, la población del bagre está fuertemente presionada por la pesquería artesanal, lo que explica la fuerte reducción en la oferta de este recurso (CCI-INCODER 2006) e implicaciones biológicas relacionadas con la reducción en su variabilidad genética debido a la reducción en el tamaño de la población lo que potencia la endogamia (Gallo y Díaz 2003).
Las hembras S. cuspicaudus y de P. magdaleniatum se caracterizan por poseer mayor longitud estándar que los machos. De acuerdo con los resultados del estudio, el tamaño de las hembras parece ser importante en la época reproductiva. Diversos autores (Wooton 1999, Moyle y Cech 2004) afirman que en especies sin cuidado parental y desoves pelágicos, la fecundidad es función del tamaño así que hembras con mayores tallas pueden repercutir en posturas más numerosas y mayor tamaño de la cohorte 0+.
]]> Las especies presentaron diferentes proporciones sexuales. Mientras que en P. magdaleniatum se observó una proporción 1:1, en S. cuspicaudus el número de hembras fue superior al de los machos pero sólo durante el segundo ciclo hidrológico intra-anual. Esta proporción encontrada es similar a la encontrada en las dos especies para toda la cuenca del río Magdalena (CCI-INCODER 2006). Diversos autores (Darwin 1871, Fisher 1929, Pianka 1982) discuten las causas del desvío del valor teórico. La proporción sexual 1:1 responde a una relación donde un individuo de una especie diploide proviene de un macho y de una hembra y las modificaciones a esta proporción se originan en la cantidad de esfuerzo invertido por los parentales en la producción de individuos de determinado sexo y en la tasa de mortalidad. Los bagres considerados en este estudio son especies dioicas con fecundación cruzada –gonocóricas– (Wooton 1999) y sin cuidado parental (Vazzoler 1996, Winemiller 1989). De esta forma, es posible que el desvío en la proporción sexual en la población de individuos adultos de S. cuspicaudus sea el resultado de las áreas donde fueron capturados y del tipo de aparejo utilizado para su captura. Aunque los pescadores no han observado segregación de hábitat según el sexo ni selectividad de los aparejos de pesca, sería importante verificar la hipótesis de que existan este tipo de factores que influyen en resultados provenientes de pesquerías artesanales.Los individuos desovantes de S. cuspicaudus y P. magdaleniatum presentaron una proporción sexual similar durante los periodos hidrológicos observados, con excepción de la dominancia de hembras en el blanquillo en el segundo ciclo hidrológico. Es importante reseñar que la información analizada proviene de capturas de pescadores y que, si bien los aparejos de pesca y los lugares de captura pueden ser particulares al sexo, la información oral proveniente de los pescadores afirma que no existe tal selectividad y ésto se observa en la proporción sexual observada; sin embargo, con la información obtenida no es posible explicar el caso de la dominancia de hembras del blanquillo durante el segundo ciclo hidrológico intra-anual.
La fecundidad de S. cuspicaudus y P. magdaleniatum es alta, los ovocitos son pequeños y su diámetro es similar. Vazzoler (1996) reportó fecundidades y diámetros similares para especies de Pseudoplatystoma y Buendía et al. (2003) para la población de S. cuspicaudus en la cuenca del río Sinú. Winemiller (1989) plantea que la estrategia reproductiva óptima en un ambiente determinado se define como la maximización o minimización de un parámetro demográfico y su relación con la tasa de cambio de los otros parámetros. Este autor retoma la propuesta de Pianka (1970) sobre estrategias de vida k y r y renombra a la estrategia k como ''equilibrio'' (especies sedentarias en ambientes relativamente estables, con ciclos de vida largos, baja fecundidad, alta inversión energética en tamaño de ovocito y en cuidado a la progenie, con varios desoves en una misma temporada reproductiva) y presenta dos variaciones de la estrategia r: r-oportunista (especies pequeñas que colonizan rápidamente ambientes perturbados, con ciclos de vida cortos, baja fecundidad, baja inversión energética en tamaño de ovocitos y cuidado de la prole y desoves frecuentes) y r-estacional (especies altamente móviles, ciclos de vida prolongados, alta fecundidad, una o dos reproducciones anuales y generalmente ajustados con los periodos hidrológicos y baja inversión energética en tamaño de ovocito y en cuidado de la prole). De acuerdo a este autor y con base en la información obtenida en este trabajo en la cuenca del río Magdalena, P. magdaleniatum y S. cuspicaudus poseen una estrategia reproductiva r-estacional puesto que presentan dos periodos reproductivos al año, alta fecundidad y ausencia de cuidado parental puesto que luego del desove, las larvas derivan en el canal del río.
La presencia de individuos con gónadas en reposo y, en particular, cuando el porcentaje de individuos desovantes se incrementa durante las aguas subiendo y altas, sugiere que parte de la población de S. cuspicaudus y P. magdaleniatum que ha migrado, no lo hace para reproducirse. Según Sumpter (1990), hay dos tipos básicos de mecanismos fisiológicos que se desarrollan durante la reproducción: los exógenos (relacionados con las señales ambientales) y los endógenos (ritmo propio al individuo). Por otro lado, Kawakami (2003) y Valderrama (1972) describen como las migraciones de algunas especies de Pimelodidae son provocadas por la migración de sus presas y no necesariamente tienen repercusiones reproductivas, pues es común observar a estas especies alimentándose de individuos de Characiformes que participan de la migración. De esta forma, es posible que la fracción de la población de estos pimelódidos tenga dos componentes: individuos que migran y se reproducen durante el primer periodo hidrológico de estiaje-crecientes y otro que lo hace en el segundo periodo. Individuos que migran y no se reproducen puede resultar del ritmo reproductivo endógeno propio y que su movilización hacia otras áreas sea únicamente de carácter trófico.
El bienestar de las poblaciones de P. magdaleniatum y S. cuspicaudus esta asociado al periodo hidrológico. Mientras que durante las crecientes el bienestar tiende a reducirse, en aguas altas se incrementa. El cambio en el factor de bienestar somático está asociado generalmente a periodos de maduración gonádica o a cambios en la intensidad alimenticia (Wooton 1999). Diversos autores (Carosfeld et al. 2003) han señalado que el comportamiento de los peces en buena parte de los ríos tropicales y subtropicales está asociado con las lluvias. Durante las crecientes, las especies invierten la energía somática disponible en tejido reproductivo y desoves y, en el final del periodo de aguas altas, recuperan la energía invertida en la pasada reproducción migrando río abajo hacia hábitats que ofrezcan alimento (migración alimentaria).
La localización de las áreas y el momento de desove de S. cuspicaudus y P. magdaleniatum son importantes para la sobrevivencia de la prole, puesto que permite su desarrollo hasta fases avanzadas y el aprovechamiento de las áreas de crianza ofrecidas en las aguas altas. Barthem y Goulding (1997) afirman que los nuevos individuos producto de la reproducción de Brachyplatystoma y Pseudoplatystoma pueden derivar distancias entre 2.500 y 3.500 km desde los lugares de desove en el piedemonte andino y que esto, basados en la velocidad del agua del río durante el periodo de aguas altas, explica la presencia de juveniles de estas especies en el estuario del río Amazonas. En la cuenca media del río Magdalena, el periodo de desove se sincronizó con las crecientes y las áreas de desove de estas dos especies se localizaron, teóricamente, entre los 131 km (S. cuspicadus) y 324 km (P. magdaleniatum) aguas arriba de la ciudad de Puerto Berrío. Esto permite que durante el tiempo que la prole deriva esta distancia, alcance la fase de flexión (momento en el que algunas aletas se encuentran desarrolladas, el ojo pigmentado y el sistema digestivo preparado para utilizar alimento exógeno) y que su ingreso a los planos de inundación, no genere altas mortalidades.
La conformación geomorfológica del plano inundable en proximidades de la ciudad de Puerto Berrío y el régimen de lluvias en el sector de Puerto Berrío pueden ser determinantes en la influencia que el río Magdalena tiene sobre su plano de inundación y por ende, en el ingreso de la prole de S. cuspicaudus y P. magdaleniatum a estos ambientes. Welcomme (1985) y Lowe-McConnell (1995) describen como las crecientes de los ríos son vitales para ofrecer a la prole de especies migratorias, los recursos (alimento y hábitat) necesarios para su sobrevivencia. Es interesante anotar que estudios realizados en el río Sinú, indican que la regulación de las crecientes que provoca la central hidroeléctrica Urra ha reducido el ingreso de larvas a las ciénagas del sistema y por lo tanto, la afectación de la oferta pesquera (CINPIC 1998). En el sector analizado de la cuenca del río Magdalena, solo durante las primeras crecientes del año se observa el acceso del río a la ciénaga La Pequeña, debido a la interacción de dos factores: ausencia de lluvias locales (lo que ocasiona reducción en la altura de la columna de agua en la ciénaga y provoca que el Caño Negro presente bajos niveles de agua) y el incremento en el nivel del agua del río Magdalena por la presencia de lluvias en los sectores altos de la cuenca. Esto permite que el río Magdalena ingrese por Caño Negro en dirección hacia la ciénaga La Pequeña sin ofrecer resistencia hidráulica importante. En el segundo periodo de crecientes, las lluvias locales no desaparecen y esto provoca que Caño Negro ofrezca resistencia a la entrada del río al plano inundable y, que en consecuencia, el ingreso de la prole durante este segundo periodo reproductivo no se observe.
Sólo en el caso de P. magdaleniatum se observó alguna relación entre la información biológica de los adultos y la densidad de sus larvas en la cuenca media del río Magdalena. Es importante tomar en consideración que las larvas fueron capturadas frente a la ciudad de Puerto Berrío y que según las distancias estimadas de deriva larval (ver Resultados), la población de adultos de S. cuspicaudus responsables de éstos desoves, probablemente no estarían siendo reclutados en la pesca de este sector de la cuenca media del río Magdalena. Así que, de manera preliminar, podemos sugerir que la utilización de indicadores biológicos provenientes de individuos adultos es válida únicamente dentro de un contexto local y que no es recomendable utilizarla como indicadora de actividad reproductiva a nivel regional. Por el contrario, el monitoreo a la densidad de larvas puede ser una herramienta más certera para definir periodos reproductivos a una escala espacial y temporal mayor pues se encuentra totalmente sincronizada con la dinámica hidrológica propia de la cuenca.
De acuerdo con el cambio en la densidad de larvas, se reconocieron dos momentos reproductivos de S. cuspicaudus y P. magdaleniatum dentro del ciclo hidrológico anual en la cuenca del río Magdalena, cada uno de ellos de menor intensidad (mayor densidad en el primer ciclo) y asociado a diferentes periodos hidrológicos. La especie S. cuspicaudus se reprodujo cuando el río tenía los menores niveles de agua mientras que P. magdaleniatum lo hizo en las crecientes. Es interesante anotar que en la cuenca del río Sinu, la reproducción de S. cuspicaudus se sucede en los periodos de crecientes (Buendia et al. 2003). Es posible que la diferenciación de los periodos de desove de estos dos pimelódidos en la cuenca del río Magdalena pueda obedecer a una estrategia relacionada con la reducción de la competencia por recursos de estas dos especies. Ésta es una hipótesis que deberá ser verificada en futuros estudios.
En conclusión, podemos afirmar que la población de P. magdaleniatum está siendo fuertemente presionada por la actividad pesquera, que las características biológicas de adultos para definir periodos reproductivos deben ser utilizadas con precaución y se aconseja utilizar la densidad de sus larvas como señal inequívoca de la ocurrencia de desoves y, finalmente, que la conectividad hidrológica entre el cauce principal y las ciénagas de su plano lateral es definitiva para permitir el ingreso del ictioplancton a estas áreas de crianza.
]]>AGRADECIMIENTOS
Esta investigación se pudo realizar gracias al apoyo de la beca 14289 del Fondo Internacional para la Ciencia (IFS) y a la colaboración en campo de los pescadores de la Asociación ASOPESCA en la ciudad de Puerto Berrio (Antioquia). El primer autor agradece de manera muy especial a los doctores Miguel Petrere Jr. y Rosseval Leite por su acompañamiento durante la realización de su trabajo doctoral.
REFERENCIAS
1. Agudelo C, Córdoba E, Salinas Y, Sánches C, Muñoz-Sosa D, Alonso JC, Arteaga M, Rodríguez O, Anzola N, Acosta L, Núñez M, Valdés H. 2000. Bagres de la Amazonia colombiana: Un recurso sin fronteras. Leticia (Colombia): Instituto Amazónico de Investigaciones Científicas, Ministerio del medio Ambiente. [ Links ]
2. Barthem R, Goulding M. 1997. The catfish connection-ecology, migration and conservations of Amazon predators. New York (USA): Columbia University Press. [ Links ]
3. Buendía D, Argumedo J, Olaya-Nieto C. 2003. Biología reproductiva del blanquillo (Sorubim cuspicaudus Littmann, Burr y Nass 2000) en el río Sinú. MVZ-Córdoba 8: 279. [ Links ]
4. Carosfeld J, Harvey B, Ross C, Baer A. 2003. Migratory fishes of South America: Biology, fisheries and conservation status. Ottawa (Canada): World Fisheries Trust. [ Links ]
5. CCI-INCODER (Corporación Colombia Internacional CCI, Instituto Colombiano de Desarrollo Rural INCODER). 2006. Informe anual de Pesca y Acuicultura Colombia. Bogotá (Colombia): Convenio CCI- INCODER. [ Links ]
6. CCI-INCODER (Corporación Colombia Internacional CCI, Instituto Colombiano de Desarrollo Rural INCODER). 2007. Informe técnico regional cuencas del Magdalena, Sinú y Atrato. Convenio Bogotá (Colombia): CCI-INCODER. [ Links ]
7. Cochran W. 1952. The x2 test of goodness of fit. Annals of Mathemathical Statistics, 23: 315-345. [ Links ]
8. Contreras P, Contreras J. 1989. Desarrollo embrionario y larval del bagre rayado Pseudoplatystoma fasciatum (Linnaeus 1776) (Pisces: Pimelodidae). Informe interno. Barrancabermeja (Colombia): INDERENA. Estación Piscícola de San Silvestre. [ Links ]
9. Darwin C. 1871. The decent of man and selection in relation to sex. New York (USA): Appleton and Company Press. [ Links ]
10. Fisher RA. 1929. The genetical theory of natural selection. Rev. 1958. New York (USA): Dover Press. [ Links ]
11. Gallo H, Diaz-Sarmiento J. 2003. Variabilidad genética del bagre rayado Pseudoplatystoma fasciatum (Pisces: Pimelodidae) en el río Magdalena (Colombia). Revista de la Academia Colombiana de Ciencias, 23 (105): 599-605. [ Links ]
12. Granado C. 2002. Ecología de Peces. Sevilla (España): Secretariado de Publicaciones, Universidad de Sevilla. [ Links ]
13. Jiménez-Segura LF. 2008. Reproductive events of migratory fish species in the middle basin of the Magdalena river. Medellín (Colombia): Final report-International Foundation for Science. [ Links ]
14. Kawakami de Resende E. 2003. Migratory fishes of the Paraguay-Parana Basin, excluding the Upper Paraná Basin. En: Carosfeld J, Harvey B, Ross C, Baer A (eds). Migratory fishes of South America: Biology, fisheries and conservation status. Ottawa (Canada): World Fisheries Trust. p. 99-156. [ Links ]
15. Kruskal W. 1952. A non-parametric test for the several simple problem. Annals of Mathematical Statistics, 12: 525-540. [ Links ]
16. Kendall Jr AW, Alhstrom EH, Moser HG. 1984. Early life history stages of fishes and their characters. En: Moser HG, Richards WJ, Cohen DM, Fahay MP, Kendall AW Jr, Richardson SL (eds). Ontogeny and Systematics of fishes. Based on international symposium dedicated to the memory of Elbert Halvor Alhstrom. Lawrence (Estados Unidos). Special publication American Society of Ichthyologist and Herpetologist, 1: 11-22. [ Links ]
17. Lowe-McConell RH. 1995. Ecological studies in tropical fish communities. Cambridge (Reino Unido): Cambridge University Press. [ Links ]
18. Mann HB, Whitney DR. 1947. On a test whether one of the two random variables is stocastically larger than the other. Annals of Mathematical Statistics, 18: 50-60. [ Links ]
19. Moyle P, Cech J. 2004. An introduction to ichthyology. Londres (Reino Unido): Prentice Hall Press. [ Links ]
20. Nakatani K, Agostinho AA, Baumgartner G, Bialetski A, Vanderlei P, Cavicchioli M, Pavanelli C. 2001. Ovos e larvas de peixes de água doce, desenvolvimento e manual de identificação. Maringá (Brasil): Universidad Estadual de Maringá, NUPELIA, Eletrobrás. [ Links ]
21. Novoa J, Cataño Y. 2005. Descripción del desarrollo embrionario y larvario del Blanquillo Sorubim cuspicaudus (Littmann, Burr y Nass, 2000) [Trabajo de pregrado en Acuicultura]. [Montería (Colombia)]: Universidad de Córdoba. [ Links ]
22. Padilla P, Alcántara B, Ismiño R. 2001. Reproducción inducida de la doncella Pseudoplatystoma fasciatum y desarrollo embrionario-larval. Folia Amazónica, 12 (1-2), IIA: 1-141. [ Links ]
23. Pianka E. 1970. On r and k selection. American Naturalist, 102: 592-597. [ Links ]
24. Pianka E. 1982. Ecología evolutiva. Barcelona (España): Ed. Omega. [ Links ]
25. Ricker WE, 1975. Computation and interpretation of biological statistics of fish population. Bulletin Fisheries Research Board of Canada, 191: 1-138. [ Links ]
26. Sumpter JP. 1990. General concepts of seasonal reproduction. En: Munro AD, Scott AP, Lam TJ (eds). Reproductive seasonality in teleosts: Environmental influences. Boca Raton (USA): CRC Press. p. 13-28. [ Links ]
27. Valderrama M. 1972. Operación subienda. Investigación pesquera. Bogotá (Colombia): Instituto de Desarrollo de los Recursos Naturales Renovables. [ Links ]
28. Valderrama-Barco M, Zárate-Villarreal M. 1989. Some ecological aspects and present state of the fishery of the Magdalena river basin, Columbia, South America. En: Dodge DP (ed). Proceedings of the international large river symposium. Canadian Special Publication Fisheries Aquatic Sciences, 106: 409-421. [ Links ]
29. Vazzoler AM. 1996. Biologia da reprodução de peixes teleósteos: Teoria e prática. Maringa (Brasil): Ed. DAUFSC. Nupelia-Universidad de Maringa. [ Links ]
30. Welcomme RL. 1985. River fisheries. Roma (Italia): FAO Fisheries Technical Paper 262. [ Links ]
31. Winemiller KO. 1989. Patterns of variation in life history among South American fishes in seasonal environments. Oecologia, 81: 225-241. [ Links ]
32. Wooton RJ. 1999. Ecology of teleost fishes. Fish and Fisheries Series 24. Londres (Reino Unido): Kluwer Academic Publishers. [ Links ]
33. Zar J. 1996. Biotatistical analysis. New Jersey (USA): Prentice Hall Press. [ Links ]
]]> Recibido: noviembre 2008;
Aceptado: mayo 2009.
]]>