Microhabitat use, diet and time of activity of four sympatric Neotropical hylid frogs (Anura: Hylidae)
JAIDER MUÑOZ-GUERRERO
VÍCTOR H. SERRANO
MARTHA PATRICIA RAMÍREZ-PINILLA
Laboratorio de Biología Reproductiva de Vertebrados, Escuela de Biología, Grupo de Estudios de la Biodiversidad, Universidad Industrial de Santander, Bucaramanga, Colombia. jaidermuz1@hotmail.com; vserrano@uis.edu.co; mpramir@uis.edu.co
RESUMEN
]]> Se estudiaron el uso del microhábitat, la dieta y la actividad reproductiva en cuatro especies simpátricas de hílidos en charcas temporales del Caribe colombiano. Se registraron ocho especies de hílidos pero sólo cuatro de ellas se estudiaron dada su abundancia y representatividad: Dendropsophus microcephalus, Scarthyla vigilans, Hypsiboas pugnax y Scinax rostratus. Durante la estación más seca del año no se encontraron ranas, así el tiempo de muestreo y las observaciones se hicieron durante las estaciones de lluvias entre 1999 y 2001. El uso del microhábitat (sustrato y altura) varió entre las especies; las especies pequeñas D. microcephalus y S. vigilans se encontraron en el agua sobre vegetación emergente, mientras que las especies de tamaños corporales mayores se hallaron en vegetación alta alrededor de los charcos (H. pugnax, S. rostratus); así, un solapamiento espacial es evidente sólo entre especies de similar tamaño. No hubo diferencias entre especies en el tipo de presas consumidas; sin embargo, hubo diferencias significativas en la frecuencia y el tamaño de las presas ingeridas en relación con el tamaño corporal: a más grandes tamaños corporales, mayores tamaños de presas consumidas. Durante las estaciones de lluvias, la abundancia y la actividad de canto de las cuatro especies no fueron continuas en el tiempo. Aunque los machos de las cuatro especies fueron reproductivamente activos todos los meses, las diferencias en el tiempo de actividad son dadas por el tiempo de actividad reproductiva de las hembras. S. vigilans y S. rostratus tienen una reproducción explosiva mientras que D. microcephalus y H. pugnax mostraron una reproducción extendida en el tiempo durante la mayor parte de la estación de lluvias. Así, se observó una segregación temporal entre las especies de tamaño corporal similar permitiendo un mínimo solapamiento en el uso de los recursos (alimento y microhábitat).Palabras clave. Uso de los recursos, dieta, Hylidae, microhábitat, actividad reproductiva.
ABSTRACT
Microhabitat use, diet, and reproductive phenology were studied in four sympatric hylid species in temporal ponds of the Colombian Caribbean. Eight hylid species were registered but only four of those were studied given their abundance and high representativeness: Dendropsophus microcephalus, Scarthyla vigilans, Hypsiboas pugnax and Scinax rostratus. During the extreme dry season frogs were not found, thus the sampling time and observations were made during rainy seasons between 1999 and 2001. Microhabitat use (substrate and height) varied among species; the small species D. microcephalus and S. vigilans were found within the pond on low submerged vegetation whereas the species of large body size were on high vegetation around the water bodies (H. pugnax, S. rostratus); then, a spatial overlap was present only between the species of similar size. There were no differences among species in the type of consumed prey; however, there were significant differences in the frequency and size of ingested prey related to body size: larger species consumed larger prey. During the rainy seasons the abundance and calling activity of the four species were not continuous in time. Although the males of the four species were reproductively active all rainy months, differences in the time of activity are related with female reproductive activity. S. vigilans and S. rostratus have an explosive reproduction where as, D. microcephalus and H. pugnax showed an extended reproductive activity during almost the whole rainy season. Thus, a temporal segregation was observed between species of similar body sizes allowing a minimal overlap in the use of the resources (food and microhabitat).
Key Words. Diet, Hylidae, microhabitat, reproductive activity, use of resources.
INTRODUCCIÓN
Se ha encontrado que existen diferencias ecológicas importantes entre las especies que coexisten en los diferentes ensambles de anuros (Lima & Magnusson, 1998; Bertoluci, 1998; Cabral & Sazima, 2000). Las especies explotan diferentes hábitats o microhábitas, recursos alimentarios, o son activas en tiempos diferentes (Zimmerman & Simberloff 1996, Moreira & Barreto 1997, Lima & Magnusson 1998). Toft (1980, 1985) ha resaltado que la forma en que los recursos espacio, dieta y tiempo son repartidos entre las especies y cómo son usados es de gran importancia para la estructuración de los ensambles de anuros. Así, las interacciones competitivas son mecanismos importantes que dan estructura a las comunidades (Albrecht & Gotelli 2001). Sin embargo, otras interacciones como depredación y mutualismo entre otras, son importantes a nivel local (Ricklefs & Schluter 1993), y a escala regional factores filogenéticos, evolutivos y eventos biogeográficos han mostrado ser también importantes en la estructura de estos ensambles (Vitt & Zani 1996, Zimmerman & Simberloff 1996).
Estudios en ensambles de especies de anuros neotropicales han demostrado que en particular las especies de hílidos constituyen un gremio importante para el estudio de comunidades, principalmente por su morfología y comportamiento, y un uso similar de los recursos de hábitat y microhábitat (Duellman 1967, Cabral & Sazima 2000). Teóricamente la competencia interespecífica por recursos limitados es más fuerte entre especies congenéricas coexistentes, debido a que ellas tienen características funcionales y morfológicas similares (Hauer & Stanford 1986). Así, es de gran interés analizar los factores que influyen en la coexistencia de especies de hílidos y la forma cómo los recursos son repartidos. Duellman (1967) y Bevier (1997) han observado que para las especies de hílidos neotropicales el solapamiento en el uso del hábitat es minimizado por diferencias en el uso particular de microhábitats en relación con el tamaño corporal. Así mismo, a nivel trófico los recursos son repartidos por tamaño de presas o frecuencia de ingestión (Duré 1999, Menéndez 2001), características también relacionadas con el tamaño corporal. Esto último principalmente se ha observado cuando las especies son estacionales y son activas en tiempos similares. Las especies de hílidos también pueden ser segregadas por su modo reproductivo, la fenología reproductiva y las estrategias de apareamiento (Donnelly & Guyer 1994).
Para probar si estas relaciones observadas entre especies de otros ensambles de anuros se establecen también entre cuatro especies simpátricas de hílidos que comparten charcos temporales o permanentes adyacentes a la ciénaga de Chilloa (Magdalena), este trabajo compara el uso de microhábitat, dieta, tiempo de actividad, y tiempo de actividad reproductiva de estas especies, determinando si existe un solapamiento en nicho ecológico definido por estos parámetros.
MATERIALES Y MÉTODOS
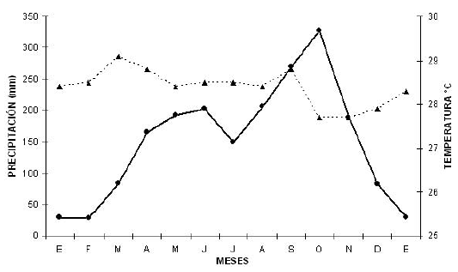
]]> Área de estudioEl área de estudio se ubica en el corregimiento de El Botillero, Municipio de El Banco, al sur del departamento del Magdalena, Colombia (09° 00’ N 73° 58’ W), a una altitud de 34 m. El área corresponde a un bosque seco tropical (bs-T) siguiendo el esquema de zonas de vida de Holdridge (1971). La temperatura media anual de la zona es de 28.3ºC, y la precipitación total anual promedio es de 1931 mm. El régimen de lluvia es bimodal tetraestacional con dos periodos de concentración de lluvias; uno de mayo a junio y otro de agosto a octubre y dos periodos secos de los cuales el más intenso se extiende de diciembre a abril y un “veranillo” (pequeña época sin lluvias) entre julio y agosto. Los máximos de precipitación se presentan en los meses de mayo y octubre; los mínimos en enero y julio. La temperatura es casi constante durante el año. La figura 1 muestra el climograma histórico de la zona para el periodo comprendido entre 1992 a 2000 (IDEAM 2002).
Figura 1. Parámetros climáticos anuales de la zona de estudio. Los datos corresponden al promedio mensual anual de precipitación (línea continua y círculos) y temperatura (línea discontinua y tríangulos). Estación climatológica IDEAM El Banco (Magdalena).
Las condiciones climáticas han sido favorables para el establecimiento de comunidades humanas haciendo parte de sistemas agroforestales de parcelas y fincas dedicadas a la ganadería intensiva, circunstancia que ha promovido la casi total desaparición de los bosques originales. La zona tiene como componente fundamental a la ciénaga de Chilloa y sus diferentes caños tributarios. La máxima altura es el cerro Botillero de 100 m que mantiene algunos remanentes de bosque. En el paisaje se observan parches aislados de bosques de galería bordeando la ciénaga de Chilloa y sus tributarios. Esta ciénaga es parte de un sistema de ciénagas conectadas con el río Magdalena en la “Depresión Momposina”. Durante las estaciones de lluvias se forman charcos temporales numerosos e irregulares, conectados o no con la ciénaga y sus drenajes.
Trabajo de campo
Los individuos se colectaron en charcos temporales porque el acceso a la orilla de la ciénaga no fue fácil durante los meses de aguas altas. De las diferentes especies de hílidos de la zona (Hypsiboas crepitans, Hyla pugnax, Dendropsophus microcephalus, D. phlebodes, Scarthyla vigilans, Scinax rostratus, Scinax ruber, Phyllomedusa venusta, Dueñez et al., 2004), cuatro especies fueron seleccionadas para este estudio debido a que fueron las especies de hílidos más abundantes en el área. Las ranas fueron colectadas durante las estaciones de lluvias ya que en un muestreo hecho durante los meses de sequía del año 2000 no fueron encontradas ni escuchadas. El estudio se realizó entonces durante diferentes meses de la estación de lluvias en tres años (septiembre de 1999, noviembre de 2000, y junio, octubre y noviembre de 2001) y durante el veranillo de mitad de año (julio, agosto de 1999). Las ranas fueron observadas y colectadas en la noche (19-22h) por dos personas, durante cinco días cada mes. Los datos de uso de microhabitat como tipo de sustrato (suelo, vegetación, tipo de plantas) y la altura de la percha (altura desde el suelo o desde la superficie del agua) fueron registradas para cada rana. Los especimenes capturados fueron eutanizados inmediatamente en etanol 10%, fijados en formol 10%, preservados en etanol 70% e ingresados a la Colección Herpetológica del Museo de Historia Natural (Universidad Industrial de Santander, UIS-A).
Trabajo de laboratorio
Todos los individuos fueron medidos con un calibrador Vernier (+ 0.02 mm): longitud rostro-cloaca (LRC) y ancho de la boca (AB). El estómago de cada espécimen fue disecado para examinar su contenido y para identificar los tipos de presas (a nivel taxonómico de Orden) y para calcular la frecuencia y el volumen de cada tipo de presa. Los diámetros mayor y menor de presas y gonadales fueron registrados [ambos diámetros se usaron para calcular el volumen de presa y gonadal, de acuerdo con Caldwell & Vitt (1999)]. El peso de los ovarios fue registrado con una balanza digital (+ 0.001 g). Para determinar el estado de espermatogénesis de los machos, los tractos reproductivos fueron post-fijados en solución de Bouin durante una hora y luego deshidratados en series ascendentes de etanol, embebidos en parafina, cortados a 6 µm y teñidos con hematoxilina y eosina. Los micropreparados así obtenidos permitieron la determinación del estado reproductivo, siguiendo la clasificación de Saidapur (1983): (1) túbulos seminíferos con espermatogonias solamente; (2) espermatocitos primarios; (3) abundantes espermatocitos primarios y secundarios, y espermátidas tempranas; (4) espermátidas transformándose en unos pocos espermatozoides; (5) espermátidas y espermatozoides abundantes; (6) espermatozoides predominantes y abundantes; y (7) espermatozoides abundantes libres, con pocos espermátides y espermatocitos. En las hembras los estados reproductivos fueron determinados macroscópicamente por la presencia de folículos yemados y oviductos convolutos. ]]> Análisis de los datos
Para determinar la existencia de diferencias significativas en el tipo de microhábitat usado por las cuatro especies fueron empleadas pruebas de G, y así como un análisis de varianza para determinar si las especies difieren en el uso del estrato vertical (altura sobre la superficie del agua o suelo). La longitud de los grupos más frecuentes de presas ingeridas por las especies fue usada para investigar si existían diferencias interespecíficas significativas en el tamaño de las presas, usando un análisis de varianza seguido por una prueba a posteriori para determinar cuáles diferían significativamente. La competencia interespecífica y el solapamiento de nicho trófico y espacial entre las especies fueron evaluados usando el índice de solapamiento de nicho según Pianka (1973). Este índice genera valores entre 0 (sin solapamiento) y 1 (solapamiento completo). Los análisis estadísticos siguieron Zar (1999) y un a = 0.05.
RESULTADOS
Tiempo de actividad
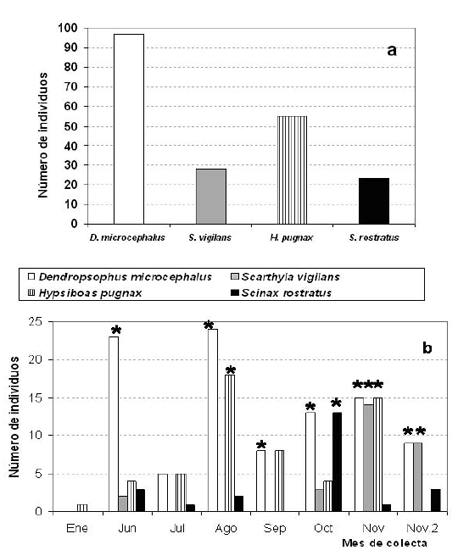
Las ranas fueron encontradas activas en las temporadas de lluvias y en el veranillo de mitad de año. Un macho no de H. pugnax fue la única rana vista durante la estación seca y por tanto los demás muestreos sólo contemplaron la búsqueda durante la estación lluviosa y el veranillo. Dos especies aparecieron casi durante todos los meses del tiempo de muestreo (D. microcephalus y H. pugnax) y ellas fueron así mismo las más abundantes, mientras que las otras dos especies fueron sólo observadas durante algunas pocas semanas (S. vigilans y S. rostratus), de manera que las cuatro especies difieren en su abundancia relativa y tiempo de actividad (Figs. 2a-b).
Figura 2. Abundancia relativa de individuos por especie. a. Número total de individuos. b. Número de individuos por especie con relación al tiempo de muestreo. El asterisco indica la presencia de hembras reproductiva.
Los machos cantores fueron decisivos para localizar y capturar las ranas durante las noches del muestreo; la mayoría de las ranas colectadas fueron estos machos y pocas hembras adyacentes a ellos; así, todos los individuos capturados fueron reproductivos. Todos los machos de las cuatro especies, incluyendo el macho de H. pugnax capturado durante la estación seca, tenían espermatozoides libres en los túbulos seminiferos y en los ductos deferentes, que fueron muy convolutos. Aunque las cuatro especies son activas y se reproducen durante la temporada lluviosa, dos especies (D. microcephalus e H. pugnax) tienen una actividad reproductiva prolongada durante varios meses, mientras que las otras dos especies S. vigilans y S. rostratus son activas y reproductivas sólo unas pocas semanas.
Uso del microhábitat
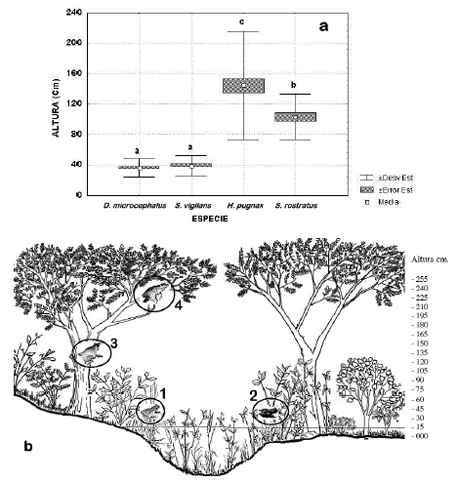
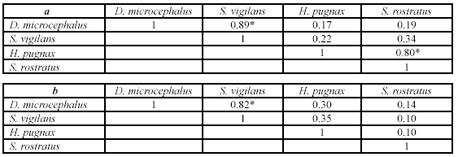
]]> Se encontraron diferencias significativas en el microhabitat usado por las especies estudiadas. D. microcephalus (G = 206.4, P < 0.001, N = 103); S. vigilans (G = 43.6, P < 0.001, N = 29); H. pugnax ( G = 196.8, P < 0.001, N = 55); S. rostratus ( G = 43.2, P < 0.001, N = 24 ). La prueba de G muestra las diferencias en la segregación por microhábitat utilizado, el valor de P es significativo para cada una de estas especies. D. microcephalus y S. vigilans, especies de pequeño tamaño, comparten el uso de vegetación herbácea emergente del nivel del agua; sin embargo, D. microcephalus se localiza principalmente en un tipo de microhábitat (pastos), mientras S. vigilans aunque usa los pastos emergentes, fue más comúnmente observada en un segundo tipo de vegetación (familia Heliconiaceae). Las especies más grandes (H. pugnax y S. rostratus) generalmente usan las ramas más altas de arbustos y árboles de la orilla y en menor frecuencia son observados en el piso de pequeñas balsas de troncos flotantes o entre las raíces emergentes de los árboles. D. microcephalus y S. vigilans no difieren en la altura de percha, ambas especies estuvieron en un rango de entre 40-50 cm por encima de la superficie del agua, pero como se mencionó arriba difieren en el tipo de vegetación preferida para perchar. En contraste, H. pugnax y S. rostratus usaron alturas de percha de rangos más amplios, entre 70 y 230 cm y 70 y 130 cm de la superficie respectivamente. Así, estas dos especies no difieren en el uso del tipo de microhábitat pero si en la altura de la percha usada (Fig. 3a), aunque pueden estar parcialmente solapadas. De esta forma, el recurso espacial alrededor de los cuerpos de agua es distribuido entre estas cuatro especies de hílidos encontrándose solapamientos entre especies de similar tamaño corporal (Fig. 3a-b, tabla 2a).Figura 3. Distribución del espacio en las charcas temporales en la zona de estudio. a. Estrato vertical utilizado por las 4 especies. Se presenta una variación significativa en la altura de percha entre las especies (ANOVA F(3. 206) = 103.46 P= 0.00, N= 210 ), las letras sobre las barras muestran las diferencias de la prueba a posteriori de Tukey. b. Distribución espacial de las especies de hílidos estudiadas. 1. Dendropsophus microcephalus, 2. Scarthyla vigilans, 3. Scinax rostratus, 4. Hypsiboas pugnax.
Tabla 2. a. Solapamiento espacial de las especies de hílidos. *Muestra los valores significativos para las especies que presentan traslape en el nicho habitacional. b. Valores de solapamiento de la frecuencia de ítems ingeridos por las especies. *valor significativo del traslape en la frecuencia de las presas.
Composición de la dieta
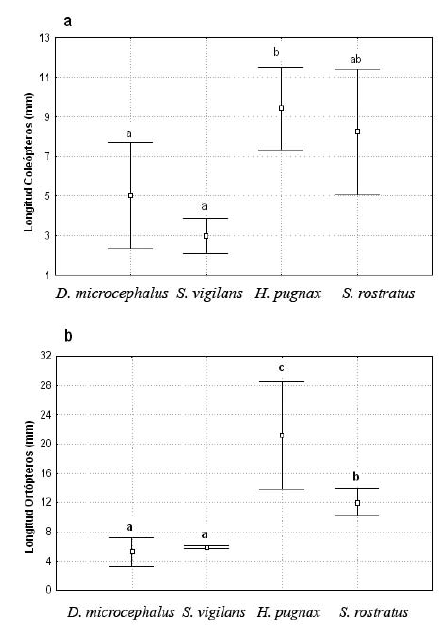
La dieta estuvo compuesta principalmente de artrópodos, y otros taxa como nemátodos, anélidos y restos de material vegetal. Los órdenes Orthoptera, Aracnida, Coleoptera, Hymenoptera y Diptera fueron los más comunes y frecuentes dentro de los contenidos estomacales, en menor proporción Blatteridae, Isoptera, Hemiptera, Homoptera y Lepidoptera (Tabla 1). Se observaron diferencias significativas entre las especies en su composición dietaria, frecuencia y tamaño de presas (Tabla 2). Las presas más voluminosas no son necesariamente las más frecuentes y viceversa. La presencia de presas raras en algunos contenidos estomacales, como Isoptera en H. pugnax y Acarii en S. rostratus, podría indicar una pequeña selección a favor de estas presas en estas especies, o su mayor disponibilidad en sus particulares microhábitats. Se encontró una variación significativa en la relación del tamaño de las presas ingeridas (Ortoptera y Coleoptera) y el tamaño corporal. Las especies de tamaño corporal mayor (H. pugnax y S. rostratus) ingirieron presas más grandes (aproximadamente entre 5 y 28 mm de longitud), mientras que las especies más pequeñas, D. microcephalus y S. vigilans, ingirieron presas más pequeñas (entre 3 y 8 mm, Fig. 4 a,b). Así, el solapamiento de la dieta es minimizado por el tamaño diferencial en las presas ingeridas entre las especies de diferentes tamaños corporales. De esta manera, las especies de tamaños similares se solapan en el tamaño de sus presas consumidas. Nosotros no evaluamos la relación intraespecífica del tamaño de la presa con el tamaño corporal ya que todos los ejemplares capturados fueron adultos de tamaños similares.
]]> Figura 4. Rangos de longitud de dos tipos de presas ingeridas por las especies. Hay una variación significativa entre los tamaños de las presas. a. Coleópteros (Kruskal-Wallis; H (3,19) = 10.16 P = 0.02) y b. Ortópteros (Kruskal-Wallis; H (3,18) = 13.96; P = 0.03). Las letras sobre las barras muestran las diferencias de la prueba a posteriori de Tukey. Se grafican los valores máximos, medios y mínimos de la longitud de las presas.
DISCUSIÓN
Las especies estudiadas son simpátricas y activas sólo durante las estaciones de lluvias. Sin embargo, su abundancia es diferente y varía en el tiempo, siendo difícil encontrar las cuatro especies abundantes al mismo tiempo. Un resultado similar fue encontrado por Rivero & Dixon (1979) y Bertoluci (1998) en diferentes ensambles de anuros donde la actividad de las especies no siempre fue sincrónica. Aunque el recurso acuático se mantiene todo el año debido a la existencia de un cuerpo de agua permanente (la ciénaga de Chilloa), las ranas no se observaron o no se escucharon durante la estación seca. Así, las lluvias y no el nivel del cuerpo de agua parecen estar directamente relacionados con la actividad y la reproducción de estas especies de ranas en esta área. De manera similar, en otras localidades neotropicales la abundancia de las especies se relaciona con la estación de lluvias, la cual favorece la reproducción de algunas de las especies durante muchos meses en esta estación (Hoogmoed & Gorzula 1979, Donnelly & Guyer 1994, Bertoluci 1998). Sin embargo, muchas especies en esos hábitats no se reproducen continuamente durante la estación de lluvias; algunas ranas tropicales se reproducen sólo por unos días cada año y sus patrones reproductivos representan un tipo de reproducción oportunista y explosiva, mientras que otras continúan reproduciéndose todo el tiempo, excepto en la estación más seca (Wells 1979, Bevier 1997).
Así, aunque los machos de nuestras cuatro especies son potencialmente reproductivos durante las estaciones de lluvias e incluso en el veranillo de mitad de año, ya que sus testículos mantienen una espermatogénesis y espermiogénesis activa, su actividad y la presencia de hembras reproductivas no fueron sincrónicas entre las especies. Un patrón de actividad más corto fue observado para S. vigilans y S. rostratus, mientras que las otras dos especies, D. microcephalus y H. pugnax, tuvieron periodos de actividad y de reproducción prolongados en varios meses de la estación lluviosa. En el mismo sentido Bevier (1997) estudió en Panamá dos especies de hílidos (Scinax ruber y Scinax boulengeri) y encontró un patrón de reproducción explosiva en S. ruber, y prolongada en S. boulengeri. Así, la distribución temporal del esfuerzo reproductivo varía entre especies congenéricas simpátricas; de acuerdo con Bevier (1997) los individuos de dos especies similares simpátricas pueden dedicar similar cantidad de energía para la reproducción, pero el tiempo en que esta actividad se realiza puede ser completamente diferente. Estas diferencias en el inicio y duración del periodo de actividad y reproducción determinan una segregación temporal en las comunidades de adultos y larvas, y pueden reducir las interacciones interespecíficas; un estudio cuantitativo más exhaustivo con datos de fenología reproductiva y tiempo de actividad para cada especie pueden probar esta hipótesis.
Las especies estudiadas se observaron activas en la noche sobre la vegetación que rodea los cuerpos de agua, difícilmente en el suelo o en la hojarasca del bosque. Estas características son compartidas por los hílidos en contraste con ranas de otras familias que componen los ensambles neotropicales de bajas altitudes como leptodactílidos, bufónidos o dendrobátidos que son más propios de microhábitats más terrestres ocupando el suelo del bosque (Lima & Magnusson 1998, Caldwell & Vitt 1999). Las especies de tamaño corporal pequeño ocupan microhábitats diferentes a los de las especies más grandes, así los tamaños corporales tienen una relación con el uso del microhábitat. Este hecho permite sugerir que el tamaño corporal favorece la segregación de especies, existiendo una mínima interacción entre las especies de tamaños corporales diferentes cuando estas son sincrónicas en tiempo de actividad y reproductivo durante la estación de lluvias. En el mismo sentido, la altura de la percha es otro factor de segregación en el caso de las especies de tamaño corporal similar. En este análisis H. pugnax y S. rostratus generalmente usan los mismos microhábitats; sin embargo, cuando estas especies se encuentran simultáneamente, ellas difieren en la altura de la percha: la primera de estas dos especies fue observada cantando en ramas de árboles más altas que la segunda. Así, una pequeña interacción es observada durante la estación de lluvias en los charcos, quizás relacionada con diferencias en microhábitats para los coros de machos y posiblemente también para la localización de las posturas. Estos hechos podrían implicar una división del recurso del charco para la coexistencia durante las temporadas de actividad y reproducción.
La aparente similitud en la composición de presas en las especies estudiadas podría estar relacionada con la disponibilidad ambiental de estos recursos. Aunque algunas especies son selectivas en su alimentación, los anfibios son generalmente considerados oportunistas ya que sus dietas reflejan la disponibilidad del tipo y tamaño de las presas (Duellman & Trueb 1986). Menéndez (2001) analizando un ensamble ecuatoriano encontró que los hílidos, contrario a otros anuros, se alimentan oportunísticamente sobre la vegetación. De manera similar Parmelee (1999) encontró que algunos hílidos tienen dietas más amplias, mientras otros parecen ser especialistas en presas grandes.
La variación en el tamaño de las presas entre especies fue marcada; las especies más grandes ingieren presas más grandes, y por tanto hubo un solapamiento significativo entre las especies de tamaños corporales similares. Duré (1999) y Menéndez (2001) encontraron que los individuos que tienen mayores tamaños corporales tienen cabezas y bocas más grandes, y por tanto pueden alimentarse de presas más grandes. El consumo energético para conseguir presas en especies grandes podría ser compensado con presas de tamaños más grandes, mientras que las especies pequeñas podrían tener que gastar más energía en la ingesta de presas más grandes (Pough et al. 1998) y estar limitados a consumir presas pequeñas. Menéndez (2001) obtuvo resultados similares entre las especies estudiadas de hílidos en su localidad. El estableció que diferencias significativas en el tamaño de las presas ingeridas en relación con su tamaño corporal, las cuales podrían explicar la repartición del nicho trófico. Sin embargo, la coexistencia de especies de hílidos podría no ser explicada solamente por la división del recurso alimenticio. En estas especies el factor más importante en la división de los recursos parece ser el uso del microhábitat ya que estas ranas son cazadoras pasivas sobre la vegetación. Así, los recursos tróficos y de microhábitat están relacionados y permiten la segregación de especies de tamaños corporales similares.
Es interesante resaltar que del par de especies pequeñas (S. vigilans y D. microcephalus) que comparten similares microhábitats, una de ellas tiene un patrón corto de actividad, mientras que la otra es activa casi toda la temporada de lluvias. De manera similar del par de especies grandes (H. pugnax y S. rostratus) que ocupan microhábitats de vegetación más altos, cada una tiene un patrón temporal de actividad diferente, siendo más prolongado el de H. pugnax. Esto podría implicar que la asincronía de actividad y de reproducción disminuye las interacciones entre especies de similares tamaños, nichos trófico y espacial, facilitando de esta forma su coexistencia. Así, en las estaciones de lluvias el grado de solapamiento en los nichos trófico y espacial es bajo debido a que no todas las especies son activas simultáneamente, y especialmente entre especies de tamaños similares, las cuales tienen diferentes patrones temporales de actividad dentro de la temporada de lluvia. Sin embargo, cuando el tiempo de actividad de cada una de las especies coincide, el recurso espacial es distribuido de tal manera que el solapamiento es disminuido entre los pares de especies de tamaños similares.
AGRADECIMIENTOS
]]> Agradecemos a los habitantes del corregimiento de El Botillero por su colaboración, especialmente a Javier Polo y su familia por la atención durante nuestra estadía. Igualmente y de manera especial a cada uno de los jóvenes que nos acompañaron en los recorridos de colecta. Al profesor John Lynch del Laboratorio de Anfibios del Instituto de Ciencias Naturales en la Universidad Nacional por su permanente colaboración. Juan P. Ramírez revisó el texto. CORPAMAG dio el permiso de colecta e investigación.LITERATURA CITADA
1. Albrecht, M. & N. J. Gotelli. 2001. Spatial and temporal niche partitioning in grassland ants. Oecologia 126: 134-141. [ Links ]
2. Bertoluci, J. 1998. Annual patterns of breeding activity in Atlantic rainforest anurans. Journal of Herpetology 32: 607- 611. [ Links ]
3. Bevier, C.R. 1997. Breeding activity and chorus tenure of two neotropical hylid frogs. Herpetologica 53: 297–311. [ Links ]
4. Cabral, P. & I. Sazima. 2000. Structure of an anuran community in a montane meadow in southeastern Brazil: effects of seasonality, habitat, and predation. Amphibia-Reptilia 21: 439-461. [ Links ]
5. Caldwell, J. P. & L. J. Vitt. 1999. Dietary asymmetry in leaf litter frogs and lizards in a transitional northern Amazonian rain forest. Oikos 84: 383-397. [ Links ]
6. Duellman, W. E. 1967. Courtship isolating mechanisms in Costa Rican hylid frogs. Herpetologica 23 : 169-183. [ Links ]
7. Duellman, W. E & L. Trueb. 1986. Biology of Amphibia. McGraw- Hill, Nueva York. [ Links ]
8. Donnelly, M.A., & C. Guyer. 1994. Patterns of reproduction and habitat use in an assemblage of Neotropical hylid frogs. Oecologia 98:291–302. [ Links ]
9. Dueñez, F., J. Muñoz & M.P. Ramírez-Pinilla. 2004. Inventario prelimirar de la herpetofauna del Corregimiento de Botillero (El Banco, Magdalena). Actualidades Biológicas 81: 64 - 75. [ Links ]
10. Duré, M.I. 1999. Interacciones en los nichos tróficos de dos especies sintópicas de la familia Hylidae (Anura) en un área subtropical de Argentina. Cuadernos de Herpetología 13: 11-18. [ Links ]
11. Hauer, F.R. & J.A. Stanford. 1986. Ecology and coexistence of two species of Brachycentrus (Trichoptera) en Rocky Mountain river. Canadian Journal of Zoology 64: 1469-1474. [ Links ]
12. Holdridge, L.R. 1971. Forest Enviroments in Tropical Life Zones: A Pilot Study. Pergamon Press, Nueva York. [ Links ]
13. Hoogmoed, M.S. & S.J. Gorzula. 1979. Checklist of the Savanna Inhabiting Frogs of the El Manteco Region with Notes on their Ecology and the Description of a New Species of Treefrog (Hylidae: Anura). Zoologishe Mededelingen Leiden 54: 183-216. [ Links ]
14. IDEAM (Instituto de Hidrología, Metereología y Estudios Ambientales). 2002. Sistema de Información Nacional Ambiental. Estación El Banco. Disponible en http://www.ideam.gov.co [ Links ]
15. Lima, A.P. & W.E. Magnusson. 1998. Partitioning seasonal time: interactions among size, foraging activity and diet in leaf-litter frogs. Oecologia 116: 259-266. [ Links ]
16. Menéndez, P.A. 2001. Ecología trófica de la comunidad de anuros del Parque Nacional Yasuní en la Amazonía Ecuatoriana (Manuscrito- Tesis de grado). Facultad de Ciencias Exactas y Naturales. Pontificia Universidad Católica del Ecuador. Quito. [ Links ]
17. Moreira, G. & L. Barreto. 1997. Seasonal variation in nocturnal calling activity of a savanna anuran comunity in central Brazil. Amphibia-Reptilia 18:49-57. [ Links ]
18. Parmelee, J.R. 1999. Trophic Ecology of a Tropical Anuran Assemblage. Scientific Papers. Natural History Museum. The University of Kansas 11:1-59. [ Links ]
19. Pianka, E. 1973. The structure of lizard communities. Annual Review of Ecology and Systematics 4: 53–74. [ Links ]
20. Pough, F.H., R.M. Andrews, J.E. Cadle, M.L. Crump, A.H. Savitzky. & K.D. Wells. 1998. Herpetology. Ed Prentice Hall. Nueva Jersey. [ Links ]
21. Ricklefs, R. E. & D. Schluter, 1993. Species Diversity in Ecological Communities. The University of Chicago Press, Chicago. [ Links ]
22. Rivero, B.C. & J. M. Dixon. 1979. Origin and Distribution of the Herpetofauna of the Dry Lowland Regions of Northern South America. Pp: 281-298 en: W.E. Duellman (Editor). 1979. The South American Herpetofauna: Its Origin, Evolution, and Dispersal. [ Links ]
23. Saidapur, S.K. 1983. Patterns of testicular activity in Indian amphibians. Indian Rew Life Sci 3:157-184 En: W.S. Huang; J. Yi lin. & J. Yu-Lin Yu (Eds). 1997. Male reproductive cycle of the toad Bufo melanostictus in Taiwan. Zoological Science 14: 497-503. [ Links ]
24. Toft, C. A. 1980. Feeding ecology of thirteen syntopic species of anurans in a seasonal tropical environment. Oecologia 45: 131- 141. [ Links ]
25. Toft, C. A. 1985. Resource partitioning in Amphibians and Reptiles. Copeia 1985: 1-21. [ Links ]
26. Vitt, L. J. & P. A. Zani. 1996. Organization of a taxonomically diverse lizard assemblage in Amazonian Ecuador. Canadian Journal of Zoology 74: 1313-1335. [ Links ]
27. Wells, K. D. 1979. Reproductive behavior and male mating success in a neotropical toad Bufo typhonius. Biotropica 11: 301-307. [ Links ]
28. Zar, J. H. 1999. Biostatistical Analysis. Printice Hall. Nueva Jersey. [ Links ]
29. Zimmerman, B. L. & D. Simberloff. 1996. An historical interpretation of habitat use by frogs in a Central Amazonian Forest. Journal of Biogeography 23: 27-46. [ Links ]
Recibido: 07/03/2007
Aceptado: 03/09/2007