A SYNOPSIS OF THE FRULLANIACEAE (MARCHANTIOPHYTA) FROM COLOMBIA
Sinopsis de la familia Frullaniaceae (Marchantiophyta) para Colombia
ROBBERT GRADSTEIN
JAIME URIBE-M.
Muséum National deHistoire Naturelle, Dept. Systématique et Evolution, UMS 7205, Case Postale 39, 57 rue Cuvier, 75231 Paris cedex 05, France.
Instituto de Ciencias Naturales, Facultad de Ciencias, Universidad Nacional de Colombia, Apartado 7495, Bogotá D.C., Colombia. juribem@bt.unal.edu.co
]]> ABSTRACTFrullania (Frullaniaceae) is the second-largest liverwort genus of Colombia (after Plagiochila), with 59 recorded species in six subgenera. In this paper keys are provided for the identification of 42 Frullania species reported from Colombia together with brief descriptions of their main morphological characters, distribution, habitat and elevation in the country, as well as worldwide distribution. Some species recorded from neighbouring countries, but not yet from Colombia, are also included in the keys. The remaining species reported from Colombia are considered doubtful taxa. Frullania apollinarii Steph. is a new synonym of F. lobato-hastata Steph.
Key words. Colombia, elevational distribution, Frullania, identification keys, liverworts, Marchantiophyta, world distribution.
RESUMEN
Frullania (Frullaniaceae) es el segundo género en número de especies de Colombia (después de Plagiochila), con 59 especies en seis subgéneros. En este artículo se proveen claves para la identificación de 42 especies de Frullania registradas para Colombia, además se incluyen breves descripciones de los caracteres morfológicos diagnósticos, distribución, hábitat y gradiente altitudinal en el país, así como la distribución mundial. Algunas especies registradas de países vecinos, pero aún no registradas para Colombia son incluidas en la clave. Las demás especies registradas para Colombia se consideran taxones dudosos. Frullania apollinari Steph. es un nuevo sinónimo de F. lobato-hastata Steph.
Palabras clave. Colombia, distribución altitudinal, Frullania, claves de identificación, hepáticas, Marchantiophyta, distribución mundial.
Recibido: 07/03/2011
Aceptado: 19/08/2011
INTRODUCTION
The liverwort family Frullaniaceae consists of two genera, Frullania with about 300-350 species worldwide and Neohattoria with one species in eastern Asia (Gradstein et al., 2001, with updates). In Colombia, Frullania is the second-largest liverwort genus (after Plagiochila) with 59 species. The first records from Colombia were by Alexander von Humboldt and Aimée Bonpland who collected Frullania atrata (Sw.) Nees (as Jungermannia atrata Sw.) and F. riojaneirensis Raddi (as Jungermannia obscura Sw.) between Popayán and Almaguer, Dept. Cauca (Kunth, 1822). Gottsche (1864) reported 17 species from various localities based on specimens collected by Alexander Lindig and by José Jeronimo Triana. The majority of these early collections from Colombia have not been re-examined critically; those studied by Gottsche were presumably destroyed in 1944 with the bombing of the herbarium of Berlin. A further 30+ species were reported as new to Colombia by Jack & Stephani (1892), Stephani (1901-1905), Herzog (1942, 1955), Bonner (1965), Robinson (1967) and others. Stotler (1969) in his revision of the neotropical species of Frullania subgenus Thyopsiella Spruce (as subg. "Frullania") accepted 10 species for Colombia and reduced several others to synonymy. Yuzawa (1991) in his world monograph of subgenus Chonanthelia Spruce recorded 19 species from Colombia.
]]> The first edition catalogue of the liverworts and hornworts of Colombia by Gradstein & Hekking (1979) listed 58 species of Frullania for the country. This number has remained almost unaltered in subsequent editions (Uribe & Gradstein, 1998; Gradstein & Uribe, in press) in spite of intensive recent collecting activities and discovery of species new to the country, such as F. kunzei (Pinzón et al., 2003) and F. dulimensis Uribe (Uribe, 2006). Thus, the second edition of the catalogue lists 58 species (Uribe & Gradstein, 1998) and the third edition 59 species (Gradstein & Uribe, in press). The lack of increase in the number of species is explained by the reduction of various species to synonymy, thus counterbalancing the new floristic records. Examples of recent new synonymy include F. atrosanguinea Tayl. (= F. peruviana; Uribe, 2008), F. crinoidea Spruce ex Steph. (= F. atrata; Uribe & Gradstein, 2003), F. mathanii Steph. (= F. peruviana; Uribe, 2008) and F. mucronata (Lehm. & Lindenb.) Lehm. & Lindenb. (= F. brasiliensis; Uribe & Gradstein, 2003).In this paper, keys are provided for the identification of 42 species of Frullania from Colombia which we have been able to recognize with certainty. A few well-defined species known from neighbouring countries but not yet from Colombia are also included in the keys. For each species from Colombia a brief characterization is given of its distribution, habitat and elevation in the country, its world distribution, as well the main diagnostic morphological characters separating the species from related ones. The remaining species recorded from Colombia are little known taxa that need more study and may turn out to be synonyms; they are listed under "Doubtful taxa" or mentioned briefly in the discussions under the species.
The species are keyed according to the six subgenera to which they are traditionally assigned (Spruce, 1884; Uribe, 2008): subg. Chonanthelia with 17 species from Colombia, subg. Diastaloba with 2 species, subg. Frullania (= Trachycolea) with 1 species, subg. Homotropantha with 1 species, subg. Meteoriopsis with 4 species and subg. Thyopsiella with 17 species. A first morphological-phylogenetic analysis of the genus Frullania (Uribe, 2008) recovered four of these subgenera as monophyletic s.l. (incl. paraphyly; Hörandl & Stuessy, 2010): subg. Frullania, subg. Homotropantha, subg. Meteoriopsis and subg. Thyopsiella. The subgenera Chonanthelia and Diastaloba, however, proved to be polyphyletic. Hentschel et al. (2009) in a molecular-phylogenetic study based on a broader sampling confirmed the polyphyly of subg. Diastaloba and the monophyly of subg. Frullania and subg. Homotropantha; in addition, subg. Chonanthelia turned out to be monophyletic in the latter study. Subgenus Thyopsiella, however, was recovered as two poorly supported clades, a "temperate" clade containing the type of subg. Thyopsiella (F. tamarisci) and a few other species, and a "tropical" clade which included the majority of the Thyopsiella species and the type of subg. Meteoriopsis (F. peruviana). Accordingly, the temperate clade was called subg. Thyopsiella and the tropical clade was called subg. Meteoriopsis. In the present treatment, however, we have retained the six subgenera of neotropical Frullania as traditionally circumscribed. The new defintions of Thyopsiella and Meteoriopsis proposed by Hentschel et al. (2009) must be considered premature due to the lack of robust support for the two clades and the very limited sampling of the subgenus Meteoriopsis. A reclassification of the polyphyletic subgenus Diastaloba, finally, should await the urgently needed systematic revision of this group, especially of the neotropical species which have not been studied critically.
DISTRIBUTION PATTERNS
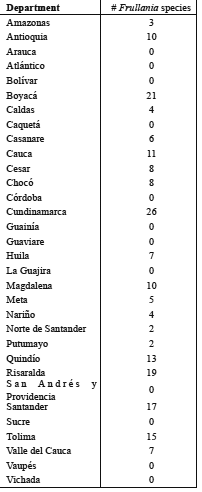
About half of the species of Frullania recorded from Colombia (22) have very wide geographical ranges and occur throughout tropical America, from Mexico to southern Brazil (see below). Six of them also occur in the Old World tropics and are pantropical species. The remaining species (20) are largely restricted to the Andes; some of these extend to the mountains of Central America and Mexico and one species (F. standaertii) occurs also in East Africa. Eight Andean species occur only in the northern Andes, from northern Peru to Venezuela (sometimes also in Costa Rica) and two species (F. bogotensis, F. dulimensis) are only known from Colombia. The geographical distribution patterns of the Colombian species of Frullania are as follows:
Pantropical: apiculata, arecae, ecklonii, ericoides, nodulosa, riojaneirensis
Neotropical: atrata, beyrichiana, brasiliensis, caulisequa, confertiloba, convoluta, cuencensis, dusenii, ecuadorensis, gibbosa, intumenscens, kunzei, macrocephala, montagnei, pittieri, setigera
Andes, sometimes extending to Central America and Mexico: grandifolia, laxiflora, paradoxa, peruviana, pluricarinata, sphaerocephala, standaertii, tetraptera, tunguraguana, winteri
Northern Andes, sometimes extending to Costa Rica: bicornistipula, clandestina, formosa, jelskii, lobato-hastata, meridana, mirabilis, ringens
Colombia only: bogotensis, dulimensis
Table 1. Number of Frullania species recorded from the Departments of Colombia
The elevational distribution of the species diversity of Frullania in Colombia is typically hump-shaped which the highest diversity (75% of the species) occurring at mid-elevations, between 1000- 2000 m (Table 2). The elevational distribution pattern of Frullania in Colombia is similar to that of liverworts in general (Uribe & Gradstein. 1999; data from 750 spp.) and apparently reflects the prevalence of epiphytism in liverworts and the preference of liverworts for humid ecological conditions. Similar hump-shaped richness patterns with peaks at mid-elevations have been reported for other plant groups, for example Melastomataceae and ferns, which like liverworts have strong ecological preferences for humid conditions (Kessler, 2000; Kluge et al., 2006). The elevation pattern of species richness detected in Frullania may thus be similar to that of many other plant groups of Colombia.
The number of Frullania species decreases to the páramo (17 spp.) but no species of Frullania is exclusive to the páramo belt even though F. decidua has been found only in páramo at 3800. The latter species occurs in Ecuador at much lower elevation in the forest belt (type location!), indicating that its distribution in Colombia is insufficiently known. Species richness is lowest in the submontane and lowland areas with only 10 and 13 species, respectively. Frullania kunzei and F. nodulosa, are only known from Amazonia below 500 m. The latter two are common and widespread lowland taxa of tropical America and F. nodulosa occurs also in tropical Africa and Asia. A third species recorded from Amazonia, F. caulisequa, is also known from the Andean region where it is reported up to 3000 m. The high-Andean records of this species should be checked, however, because elsewhere in tropical America F. caulisequa is a lowland or lower montane species (Gradstein & Costa, 2003; León-Yánez et al., 2006). Possibly, confusion with some other member of the subgenus Diastaloba to which F. caulisequa belongs has occurred. Such confusion may well have happened since the neotropical species of subg. Diastaloba have not been revised and are still very poorly known.
Table 2. Altitudinal distribution of Colombian Frullania.
KEYS AND DESCRIPTIONS
Frullania Raddi
Plants small to large, reddish-green to reddish-brown or purplish, sometimes pure green, creeping, ascending or pendent, regularly or irregularly 1-3-pinnately branched. Branches Frullania-type; innovations normally lacking. Stems rigid, made up of thick-walled cells. Leaves incubous with a very short, subtransverse insertion, divided into a large dorsal lobe, a smaller ventral lobule, and a small stylus between lobule and stem. Leaf lobe usually ovate-orbicular, apex rounded to acute to acuminate, margins entire, rarely toothed. Leaf lobule almost free from the dorsal lobe, rounded to elongate, transformed into a sac or flattened, or both. Stylus usually linear and minute, rarely large, foliar. Cells isodiametrical to (more often) elongate, usually with trigones and intermediate thickenings, the trigones often confluent and cell walls often irregularly undulate-sinuose, cuticle smooth; oil bodies finely granular; ocelli sometimes present. Underleaves small or large, 2-lobed, rarely undivided. Rhizoids in bundles from underleaf bases. Dioicous or autoicous, rarely paroicous. Androecia usually on a short, globose male branch with a few pairs of bracts and bracteoles. Gynoecia on short or elongate shoots, each with 2-5 archegonia. Perianths flattened or inflated, with 0-14 keels, the mouth contracted into a beak. Seta of numerous rows of cells. Foot of the sporophyte not penetrating into the stem. Capsule globose, wall 2-layered. Elaters attached to the capsule valves, arranged vertically inside the capsule, with 2-3-spirals. Spores large, multicellular, germination endosporic. Vegetative reproduction rare, by caducous or fragmenting leaves.
Keys to the species of Frullania from Colombia
Some species recorded from Ecuador and/or Venezuela that may occur in Colombia but are not yet known from the country are also included in the key [in brackets].
1. Introductory Key
1. Lobules distant from the stem, space between stem and lobule wider than the width of the lobule. Plants very small, creeping ............Key 2 (subg. Diastaloba)
1. Lobules close to the stem, space between stem and lobule narrower than the width of the lobule. Plants small or large ..............2
2. Lobules pendent, opening of the lobule positioned toward the stem or the apex of the plant. Amazonian lowlands (subg. Homotropantha) .....................F. nodulosa ]]>
Key 2. Frullania subgenus Diastaloba
1. Leaf apex always rounded. Plants creeping, regularly or irregularly branched ...............................................................F. caulisequa
1. Leaf apex at least in some leaves apiculate. Plants regularly (bi)pinnate, ascending from the substrate ............................................2 ]]>
Key 3. Frullania subgenus Chonanthelia
1. Lobules on the main stem with a very long laminate (= flattened) portion, which extends downwards beyond the ventral margin of the leaf lobe (sect. Cladocarpicae) .........2
1. Lobules on the main stem with a short laminate portion, which does not extend beyond the ventral margin of the leaf lobe (sect. Chonanthelia) ................................9
2. Margins of leaves and underleaves strongly undulate. Perianth 8-10-keeled .......F. arecae
2. Margins not undulate, plane. Perianth 4-keeled or 8-10-keeled ..............................3
3. Stylus large, more than 0.3 mm long, suborbicular. Perianth 4-keeled ..............................................................F. confertiloba
3. Stylus small, less than 0.1 mm long, filiform ..............................................................4
4. Laminate portion of the lobule toothed. Perianth 8-10-keeled ..................F. ringens ]]>
Key 4. Frullania subgenus Meteoriopsis
1. Leaf margins toothed ...........F. dulimensis
1. Leaf margins entire .................................2
2. Leaf apex rounded to obtuse. Female bracts entire (rarely toothed: F. grandifolia) .....3
2. Leaf apex acute to acuminate. Female bracts toothed .....................................................4
3. Underleaves ovate-oblong, 1.4-2 x longer than wide, not overlapping. Female bracts entire. Perianth distinctly exserted beyond the bracts ...............................F. convoluta
3. Underleaves suborbicular, 1-1.3 x longer than wide, overlapping. Female bracts toothed. Perianth not exserted beyond the bracts ...................................F. grandifolia
4. Underleaf base without auricles. Underleaves narrow elongate, at least 2 x longer than wide ..........................F. peruviana
4. Underleaf base with very large auricles (more than 0.15 mm long). Underleaves less than 2 x longer than wide. Known from Ecuador and Venezuela; not yet recorded from Colombia ..........................................................[F. phalangiflora Steph.]
1. Leaf apex very long and finely acuminate, the acuminate portion to 1 mm long. Known from Ecuador; not yet recorded from Colombia ....................[F. aculeata Taylor]
1. Leaf apex not very long and finely acuminate ..................................................................2
2. Lobules short, 1-1.5 × longer than wide. Plants very small, 0.5-1 mm wide, tightly prostrate. Cells in the upper part of the leaf lobes isodiametrical, evenly thickened, without trigones. Dorsal leaf base not auriculate ....................................F. kunzei
2. Lobules 1.5-3 × longer than wide. Plants larger. Cells in the upper part of the leaf lobes longer than wide, with trigones. Dorsal leaf base auriculate ......................3
3. Leaf apex apiculate to acuminate ...........4
3. Leaf apex rounded to subacute (never apiculate to acuminate) .........................13
4. Underleaves reniform (much wider than long), underleaf apex broadly truncate and with 2 small, subulate teeth ............................................................F. bicornistipula
4. Underleaves rounded, ovate to oblong, not reniform; underleaf apex not broadly truncate with 2 small teeth ......................5
5. Underleaf apices long acuminate. Lamina of first branch underleaf undivided, long acuminate .................................F. setigera ]]>
Alphabetical treatment of the Frullania species of Colombia
1. Frullania apiculata (Reinw. et al.) Nees
On bark of trees in lowland and montane rain forest, from sea level to 3000 m. Recorded from Chocó, Quindío and Santander. World distribution: pantropical.
Frullania apiculata (subg. Diastaloba) is recognized by the regularly (bi)pinnate branching and the apiculate leaf apex. The species is common throughout the tropics and has been described and illustrated in detail by Hattori (1973, 1980) for Asia and by VandenBerghen (1976) for Africa. Frullania exilis, described from the Neotropics and recorded from Colombia, has been considered a synonym of F. apiculata (e.g. Gradstein & Ilkiu-Borges, 2009) but this synonymy was not based on examination of type materials and needs confirmation.
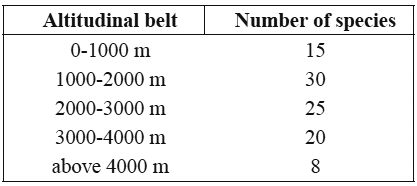
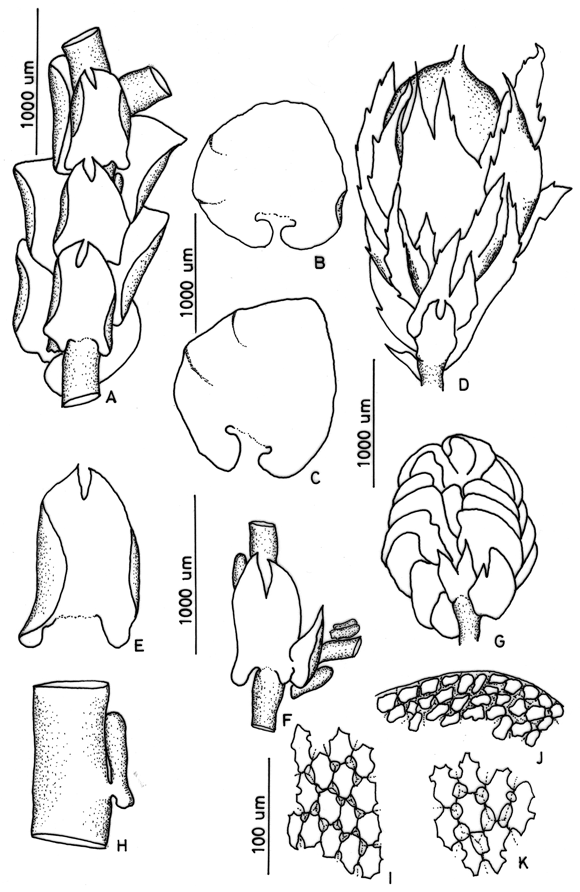
2. Frullania arecae (Spreng.) Gottsche Fig. 1
]]> On bark and rock, 1600-3400 m. Recorded from Boyacá, Casanare, Cauca, Cesar, Cundinamarca, Huila, Magdalena, Quindío, Risaralda, Santander and Tolima. World distribution: pantropical.
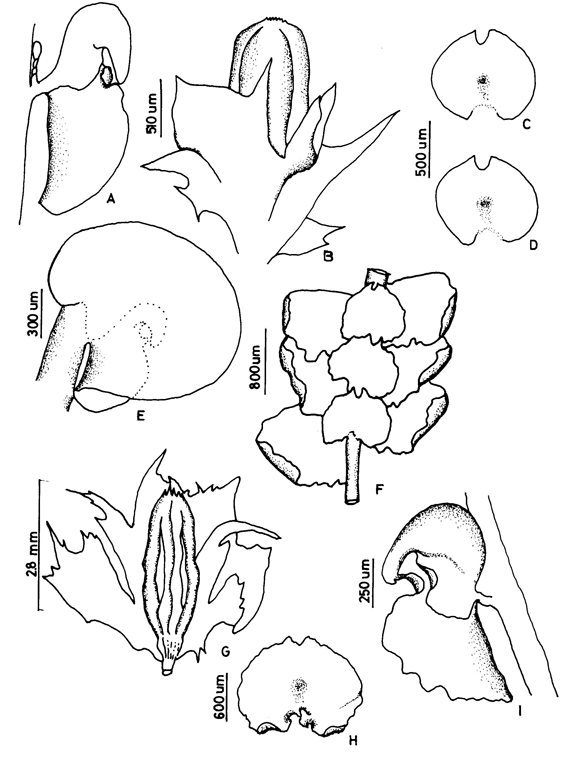
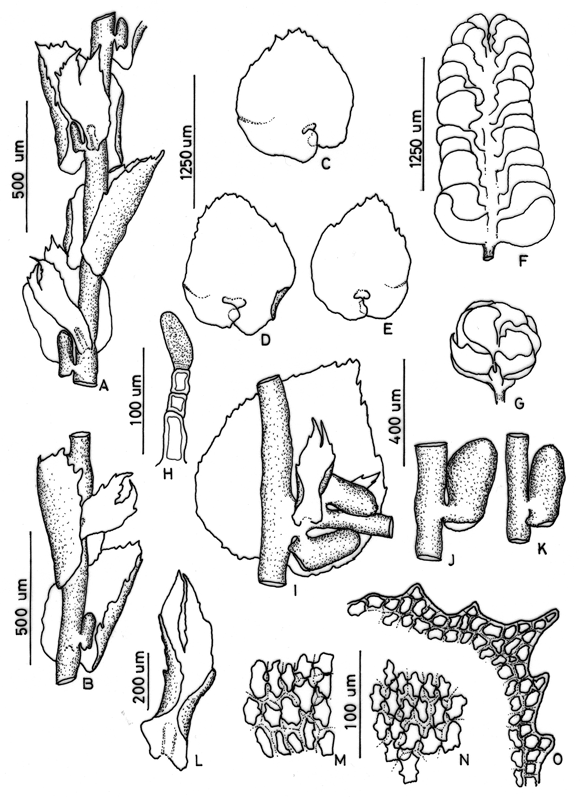
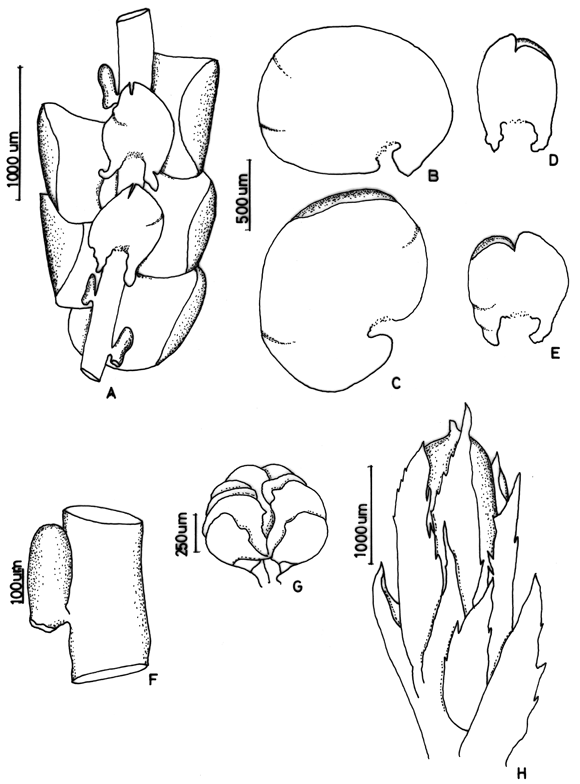
3. Frullania atrata (Sw.) Nees Fig. 2
Frullania crinoidea Spruce ex Steph. (syn. fide Uribe & Gradstein, 2003)
On bark of trees in montane rain forests, 600-3000(-4000) m. Recorded from Antioquia, Boyacá, Casanare, Cauca, Cesar, Cundinamarca, Huila, Magdalena, Meta, Santander and Tolima. World distribution: tropical America.
Frullania atrata (subg. Thyopsiella) has often been misidentified and confused with members of the subgenus Meteoriopsis such as F. convoluta and F. peruviana. However, Uribe & Gradstein (2003) showed that F. atrata is a member of the subgenus Thyopsiella and possibly related to F. intumescens, which differs by strongly recurved leaf apices (plane in F. atrata). Frullania atrata is also close to F. beyrichiana, which differs from F. atrata by the plane underleaf margins (recurved in F. atrata). The relationships of F. atrata to other members of the subgenus Thyopsiella remain poorly known and need more study.
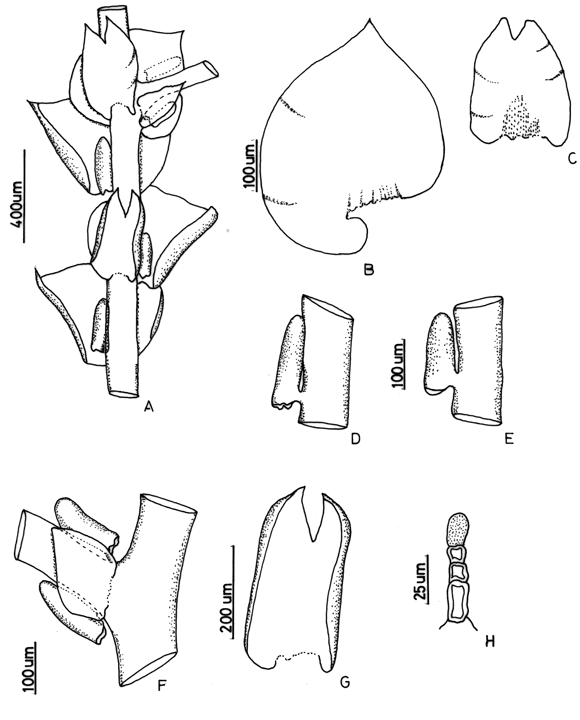
Figure 2. Frullania atrata (Sw.) Nees. A. Part of shoot, ventral view. B. Stem leaf. C. Underleaf. D and E. Leaf lobules. F. Initial branching appendages. G. Underleaf. H. Stylus. Reproduced from Cryptogamie, Bryologie with permission
On bark of trees, 1800-2100 m. Recorded from Antioquia and Cundinamarca. World distribution: tropical America.
Frullania beyrichiana (subg. Thyopsiella) is very similar to F. atrata (see under the latter species). Frullania breuteliana, which has been reported from Colombia, may be a synonym of F. beyrichiana and seems to differ from F. beyrichiana only in the slightly longer leaftip and the somewhat toothed female bracts (entire in F. beyrichiana; see Stotler, 1969).
5. Frullania bicornistipula Spruce
On bark of trees and soil, 1500-2300 m. Recorded from Cauca, Huila, Norte de Santander and Risaralda. World distribution: northern Andes and Costa Rica.
Frullania bicornistipula (subg. Thyopsiella) is a very characteristic species which is immediately recognized by the reniform underleaves with a broad truncate apex bearing 2 short, subulate teeth. The two small teeth are standing upwards and separated from each other by some distance. The long filiform stylus, 6-12 cells long, is another characteristic feature of Frullania bicornistipula.
By the long mucronate leaf apex Frullania bicornistipula is similar to F. pittieri but the peculiar underleaves of F. bicornistipula readily separate the two species.
On logs, 700 m. Recorded from Boyacá, Cundinamarca and Santander. World distribution: only known from Colombia.
Frullania bogotensis (subg. Chonanthelia) is closely related to F. riojaneirensis but differs from the latter species by the laminate portion of the lobule being broader than the sac (narrower than the sac in F. riojaneirensis) and having a folded margin (Yuzawa, 1991).
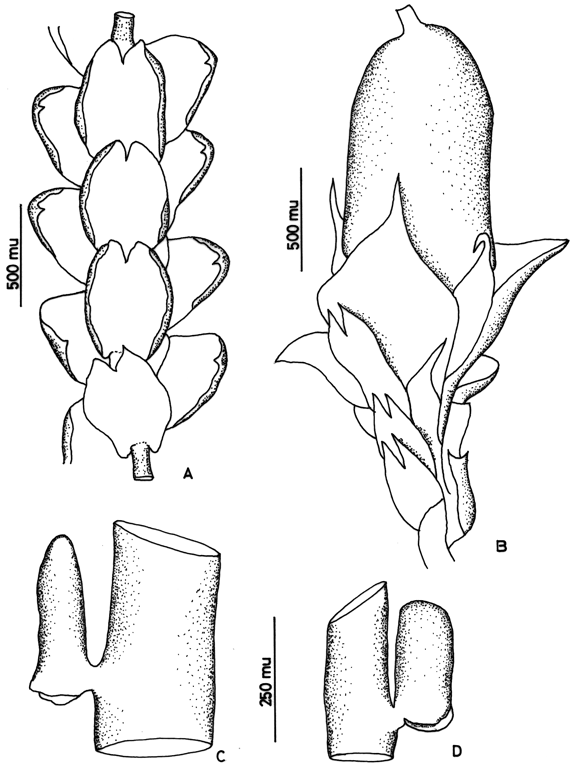
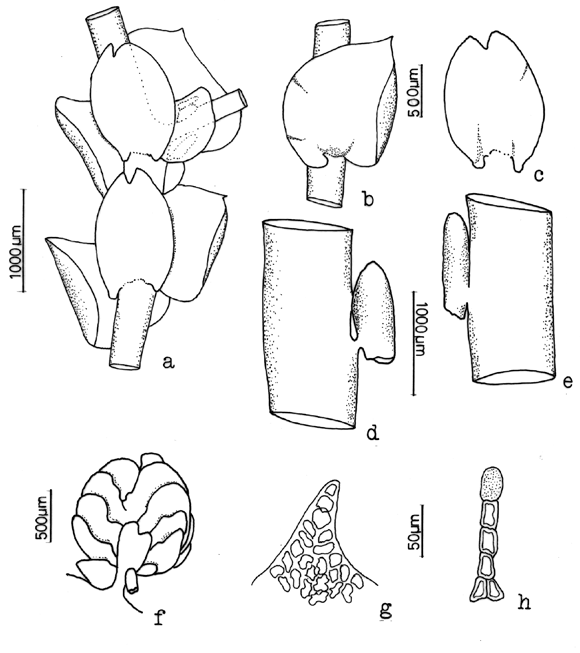
7. Frullania brasiliensis Raddi Fig. 3
Frullania mucronata (Lehm. & Lindenb.) Lehm. & Lindenb. (syn. fide Uribe & Gradstein, 2003)
Common on bark of trees, logs, soil, or rock, 1200-3900 m. Recorded from Antioquia, Boyacá, Caldas, Cauca, Cesar, Chocó, Cundinamarca, Huila, Magdalena, Meta, Nariño, Quindío, Risaralda, Santander, Tolima and Valle del Cauca. World distribution: common and widespread in tropical America.
Frullania brasiliensis is the most common neotropical species of subg. Thyopsiella and is recognized by the recurved, apiculate leaf apices and the terete perianth. Moreover, the underleaf margins are always recurved. According to Stotler (1969), F. brasiliensis has deeply bifid underleaves (to 1/3), however in material from Colombia and Ecuador of F. brasiliensis the underleaf incision varies considerably in depth (1/5-1/3).
]]>
Figure 3. Frullania brasiliensis Raddi. A. Part of shoot, ventral view. B. Perianth. C and D. Leaf lobules. Reproduced from Cryptogamie, Bryologie with permission.8. Frullania caulisequa (Nees) Nees
Frullania gymnotis Mont.
Frullania obcordata (Lehm. & Lindenb.) Lehm. & Lindenb.
On bark and rotten wood in lowland and montane rainforest areas and woodlands, from sealevel to about 3000 m. Recorded from Amazonas, Boyacá, Casanare, Cesar, Magdalena, Quindío, Risaralda, Santander and Tolima. World distribution: tropical America.
Frullania caulisequa (subg. Diastaloba) is related to F. apiculata but is immediately separated from the latter by the sparse, irregular branching and rounded leaftips. The species occurs on bark of trees at low elevations. The species was treated by Schuster (1992) under the names Frullania obcordata (Lehm. & Lindenb.) Lehm. & Lindenb. and Frullania gymnotis Mont., which are synonyms (see Gradstein & Costa, 2003). Frullania gymnotis is a form of F. caulisequa with toothed female bracts.
9. Frullania clandestina (Nees & Mont.) Nees
On bark of trees, 1500 m. Recorded from Cauca. World distribution: northern Andes (Colombia, Ecuador, Peru).
Frullania clandestina (subg. Thyopsiella) is a rare species from lower montane elevation in the northern Andes. The species is recognized by the rounded leaf apex and the large underleaves which are about as large as the leaf lobes and imbricate. By its large underleaves, F. clandestina resembles F. formosa Spruce but in the latter species the leaf apex is apiculate.
10. Frullania confertiloba Steph. Fig. 4
]]> On bark of trees and logs, 700-1700 m. Recorded from Boyacá and Santander. World distribution: scattered in tropical America.
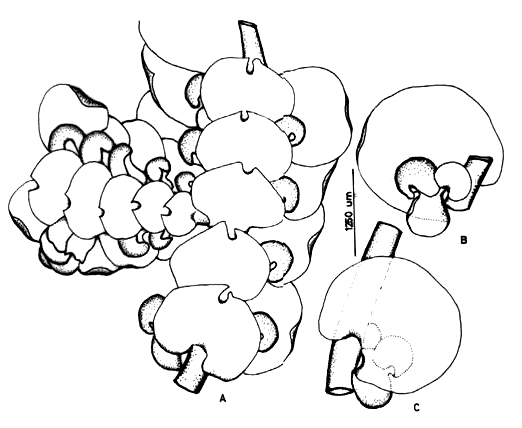
11. Frullania convoluta Lindenb. & Hampe Fig. 5
On bark of trees, pendent from branches, in upper montane cloud forest, to 3400 m. Recorded from Boyacá, Cundinamarca, Meta, Nariño, Norte de Santander, Putumayo, Risaralda, Santander and Tolima. World distribution: Central America, Dominican Republic, tropical Andes, Guayana Highlands.
Frullania convoluta (subg. Meteoriopsis) is a common species of the subgenus Meteriopsis and is recognized by pendent growth, rounded (to obtuse) leaf apex and the rather distant, ovate-oblong underleaves (Uribe, 2008). The species closely resembles Frullania grandifolia, which is the only other member of subgenus Meteriopsis with a rounded leaf apex; for differences between the two species see the key. Frullania convoluta has sometimes been confused with F. peruviana, which is also widespread in tropical America, but in the latter species the leaf apices are acute.
]]>
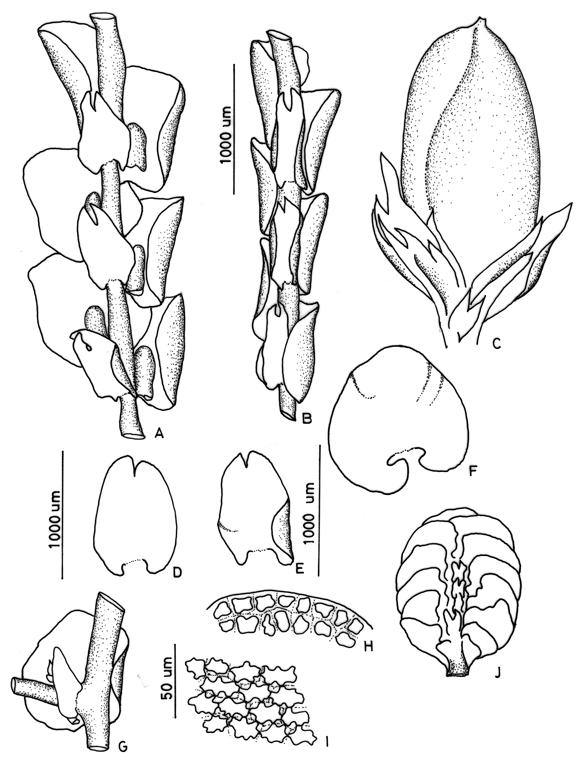
12. Frullania cuencensis Tayl.
On bark and logs, to 3000 m. Recorded from Cundinamarca and Santander. World distribution: tropical America.
Frullania cuencensis (subg. Chonanthelia) is recognized by the beaked lobule with a large sac and short laminate part, and rather deeply bifid underleaves (to 1/3) which are 2-3 times wider than the stem. The species has been confused with F. tetraptera, but the latter species has narrower and less deeply bifid underleaves and occurs at higher elevation.
13. Frullania dulimensis Uribe Fig. 6
On bark of isolated trees in open areas in the upper montane belt, growing together with other pendent species of the subgenus Meteriopsis (F. peruviana, F. convoluta), 3200 m. Recorded from Tolima (Uribe, 2006). World distribution: only known from the type specimen from Colombia.
Frullania dulimensis (subg. Meteoriopsis) is readily separated from the other members of the subgenus Meteoriopsis by the toothed leaves. The species is only known from a single collection from Colombia.
]]>
14. Frullania dusenii Steph.
On bark and rock, ca. 1700 m. Recorded from Cesar, Cundinamarca and Magdalena. World distribution: tropical America.
Frullania dusenii (subg. Chonanthelia) is recognized by the saccate lobule with a very short lamina, a narrow mouth (ca. half the width of the sac) and a plicate surface, and the rather large, rounded, imbricate underleaves which are 3-4 times wider than the stem. According to Yuzawa (1991), the species is largely restricted to elevations above 2000 m. Frullania dusenii may be confused with F. gibbosa but the latter species has squarrose leaves and a large, leaf-like stylus, and occurs at lower elevations.
15. Frullania ecklonii (Spreng. ) Gottsche et al.
On bark and rock at high elevations, 3000-3500 m. Recorded from Boyacá, Cundinamarca and Tolima. World distribution: pantropical.
Frullania ecklonii (subg. Chonanthelia) is recognized by the 10-keeled perianth, the smooth margins of leaves, lobules and underleaves, and the absence of a well-developed lobule beak. The species has sometimes been treated as a synonym of Frullania arecae but in the latter species the leaf and underleaf margins are undulate and the lobule is distinctly beaked (Yuzawa, 1991).
16. Frullania ecuadorensis Steph.
]]> On bark of trees and soil, 700-1200 m. Recorded from Boyacá and Risaralda. World distribution: scattered in tropical America.17. Frullania ericoides (Nees) Mont.
Common on bark and rock in rather dry, open woodlands, from sea level to 2800 m. Recorded from Antioquia, Boyacá, Casanare, Chocó, Cundinamarca, Quindío, Risaralda, Santander and Tolima. World distribution: pantropical.
Frullania ericoides (subg. Frullania) is one of the most drought-tolerant epiphytic liverworts and one of the few leafy liverwort species that occur in the arid lowland belt of the Galapagos Islands (Gradstein, 2008). The species is readily recognized by the squarrose and rather brittle leaves, which reproduce asexually by leaf fragmentation. The lobules are highly variable and may be saccate in some populations and laminate, without formation of watersac, in others. Frullania ericoides may be confused with Frullania gibbosa which also has squarrose leaves and occurs in the same habitat. The two species can be readily separated by their different sizes, F. gibbosa being a larger plant with leafy stems more than 2 mm wide (1-1.5 mm in F. ericoides). Moreover, the leaves in F. gibbosa are not brittle, the lobules are more broadly attached to the lobe, the perianth surfaces are smooth and the stylus is large, leaf-like (small subulate in F. ericoides).
18. Frullania formosa Spruce
On branches of trees in lowland rain forest at the foot of the Andes, ca. 500 m. Recorded from Risaralda. World distribution: only known from Colombia and Ecuador.
Frullania formosa (subg. Thyopsiella) is recognized by the flat and short apiculate leaf apex, and the large, imbricate underleaves, which are more than 3 times as wide as the stem (Stotler, 1969). By the characters of the underleaves F. formosa is similar to F. clandestina but in the latter species the leaf apex is rounded, never apiculate.
19. Frullania gibbosa Nees
Common on bark and rock, usually in rather dry, open places, 500-3200 m. Recorded from Antioquia, Boyacá, Caldas, Casanare, Cauca, Cesar, Cundinamarca, Magdalena, Risaralda, Santander and Tolima. World distribution: widespread in tropical America. ]]>
Frullania gibbosa (subg. Chonanthelia) differs from all other members of the subgenus Chonanthelia by the squarrose leaves and the large, foliose stylus. The stylus is usually covered the large and densely imbricate (and shallowly bifid) underleaves, and can be observed only by removing the underleaves. Frullania gibbosa grows in rather open locations at low elevations, often together with F. ericoides with which it may be confused. For differences between the two see under F. ericoides.20. Frullania grandifolia Steph. Fig. 7
On bark in upper montane forest, 3400 m. Recorded from Tolima (Uribe, 2008). World distribution: rare species, known from a few localities in Central America (Mexico, Costa Rica) and the northern Andes (Colombia, Ecuador).
Frullania grandifolia (subg. Meteoriopsis) is recognized by the pendent growth, the rounded leaf apex, the large, imbricate, ± rounded underleaves, and the toothed female bracts. The species is closely related to F. convoluta but in the latter the underleaves are distinctly longer than wide and not imbricate, and the margins of the female bracts are entire (Uribe, 2008).
21. Frullania intumescens (Lehm. & Lindenb.) Lehm. & Lindenb.
On bark and rock in montane rain forest areas and páramo, 1000-3800 m. Recorded from Antioquia, Boyacá, Cauca, Cesar, Chocó, Cundinamarca, Huila, Magdalena, Quindío, Risaralda, Santander, Tolima and Valle del Cauca. World distribution: tropical America. ]]>
Frullania intumescens (subg. Thyopsiella) is similar to F. brasiliensis but differs by the perianth with 3 sharp keels (terete in F. brasiliensis). Yuzawa & Koike (1989), in addition, found that the two species differ by the presence of a small foliose appendage at the base of the stylus in F. intumescens (Yuzawa & Koike, 1989). We have studied the type material of F. intumescens (Jamaica, Bancroft s.n., ex hb. Hooker 1833, lectotype in hb. S [no. B27792]) and can confirm the observations of Yuzawa & Koike (1989). However, we have not observed this appendage in any Colombian specimen identified as F. intumescens.This rare species was recorded from Colombia by Yuzawa (1991) without further information on habitat and distribution. World distribution: only known from Peru (type) and Colombia.
Frullania jelskii (subg. Chonanthelia) is characterized by the large saccate lobule with a very short lamina, the broadly recurved leaf apex, and the large underleaves (ca. 4 times as wide as the stem). The species is closely related to F. dusenii but the latter species is readily separated from F. jelskii by the narrow mouth of the sac, rounded underleaves (slightly longer than wide in F. jelskii), and the plicate lobule surface (Yuzawa, 1991).
23. Frullania kunzei (Lehm. & Lindenb.) Lehm. & Lindenb.
Frullania neesii Lindenb. (fide Gradstein & Costa, 2003)
On bark in rather open lowland woodlands and isolated trees, also on rock, 200 m. Recorded from Amazonas. World distribution: tropical America.
Frullania kunzei (subg. Thyopsiella) is readily recognized by the very small plants, less than 1 mm wide, which are creeping on bark or rock, sometimes over larger bryophytes or lichens, and always tighly appressed to the subtrate. The rather short lobules (less than 1.5 times longer than wide), the isodiametrical, evenly thickened upper leaf cells and the total absence of auricles at leaf bases, separate F. kunzei from other neotropical members of the subgenus Thyopsiella (Schuster, 1992). The species may be confused with members of the subgenus Diastaloba (e.g. F. caulisequa) which resemble F. kunzei by small size and prostrate growth, but the very different lobule in the latter species (positioned at some distance from the stem, not close to the stem) readily distinguishes them from F. kunzei. ]]>
24. Frullania laxiflora Spruce
On bark in montane rain forest areas, 1500-3500 m. Recorded from Cauca and Tolima. World distribution: scattered in the tropical Andes (Bolivia to Venezuela) and the Galapagos Islands.
Frullania laxiflora (subg. Chonanthelia) is characterized by the strongly concave leaves, the rather long laminate portion of the lobule, which is about as long as the sac yet does not extend beyond the ventral lobe margin, the decurrent beak of the lobule, and the broad, rounded underleaves with large basal auricles (Yuzawa, 1991). The species may be confused with F. riojaneirensis but in the latter species the laminate portion of the lobule extends beyond the ventral lobe margin and the beak is not decurrent.
25. Frullania lobato-hastata Steph. Fig. 8
Frullania apollinarii Steph., syn. nov.
On bark, 1800-3400 m. Colombia: Cundinamarca, Quindio and Santander. World distribution: northern Andes (Colombia, Ecuador).
Frullania lobato-hastata (subg. Thyopsiella) is a robust plant from high elevation in the northern Andes. The species is recognized by the very long, undulate auricules at underleaf bases, which are at least half as long as the underleaf lamina (lamina length measured from the middle of the underleaf insertion line to apex). The leaf apex is broadly rounded or, occasionally, short apiculate. The species resembles F. brasiliensis in the almost terete perianth but the leaf apex in F. brasiliensis is always apiculate and the underleaf auricles are small and not undulate.
Frullania apollinarii Steph., published by Stephani in the same volume of the Species Hepaticarum as F. lobato-hastata but 3 month later, is a synonym of the latter species (see Uribe, 2004a for description and illustration of F. apollinarii).
]]>
26. Frullania macrocephala (Lehm. & Lindenb.) Lehm. & Lindenb.
On bark of trees and rocks, 1400-2900 m. Recorded from Boyacá, Cundinamarca, Quindío, Risaralda and Santander. World distribution: tropical America.
Frullania macrocephala (subg. Thyopsiella) closely resembles F. brasiliensis but differs by the ± flat leaf apex (recurved in F. brasiliensis; see Stotler, 1969).
27. Frullania meridana Steph. Fig. 9
On bark of trees, 1500-3200 m. Recorded from Cundinamarca, Risaralda and Tolima. World distribution: northern Andes (Venezuela (type), Colombia).
Frullania meridana (subg. Thyopsiella) is characterized by the characters given in the key. The species is known from only few collections, and only from male plants (Uribe, 2004b).
]]>
28. Frullania mirabilis Jack & Steph.
On bark of trees and soil, 1200-2400 m. Recorded from Antioquia, Cauca, Chocó, Meta, Nariño and Valle. World distribution: northern Andes, Costa Rica.
Frullania mirabilis (subg. Thyopsiella) is easily recognized by the two long appendices at the ventral base of the leaf, one standing up and one hanging down. The species has also been described under the name F. pendulostyla Steph. (Stotler, 1969)
29. Frullania montagnei Gottsche
On bark, 1500-2100 m. Recorded from Magdalena, Santander and Tolima. World distribution: tropical America.
Frullania montagnei (subg. Thyopsiella) resembles F. paradoxa by the rounded leaf apex and the rather narrow underleaves which are less than 3 times as wide as the stem, but differs by underleaves being imbricate and having distinctly recurved margins, the undivided lamina of the first branch underleaf, and the entire margins of female bracts and bracteoles (see Key).
30. Frullania nodulosa (Reinw. et al.) Gottsche et al. ]]>
31. Frullania paradoxa Lehm. & Lindenb.
On bark of trees, 3000-3700 m. Recorded from Boyacá, Cauca, Cundinamarca, Quindío, Risaralda and Valle. World distribution: tropical Andes.
Frullania paradoxa (subg. Thyopsiella) is characterized by the rounded leaf apex, the longer than wide and rather narrow underleaves which are hardly wider than the stem, the shallowly bifid lamina (to ¼) of the first branch underleaf, and the sharply 3-keeled perianth surrounded by bracts and bracteoles with ciliate-laciniate margins (Stotler, 1969). Frullania flexicaulis Spruce, recorded from Colombia, is very similar to F. paradoxa and may be a synonym of the latter. The species differs from F. paradoxa by the female bract lobes with entire margins (usually toothed in F. paradoxa) and the somewhat more deeply bifid lamina (to ½) of the first branch underleaf.
32. Frullania peruviana Gottsche Fig. 10
Frullania mathanii Steph. (syn. fide Uribe, 2008)
Frullania atrata sensu Spruce (non typus) (fide Uribe, 2008)
On bark of tree branches in montane cloud forest areas, 1800-3800 m. Recorded from Antioquia, Boyacá, Caldas, Chocó, Cundinamarca, Magdalena, Nariño, Quindío, Risaralda, Tolima and Valle. World distribution: Central America, tropical Andes, Galapagos Islands. ]]>
33. Frullania pittieri Steph.
Frullania mucronata auct. (non typus) (fide Uribe & Gradstein, 2003)
On bark of trees, occasionally on shaded, wet rock, from sea level to 2500 m. Recorded from Boyacá, Cauca, Chocó, Quindío and Risaralda. World distribution: tropical America.
Frullania pittieri (subg. Thyopsiella) is recognized by the flat and longly mucronate to acuminate leaf apices (Stotler, 1969). The species has sometimes been misidentified as F. mucronata, which is a synonym ofF. brasiliensis (Uribe & Gradstein, 2003).
34. Frullania pluricarinata Gott.
]]> On bark of trees, 1700-3000 m. Recorded from Cundinamarca and Magdalena. World distribution: scattered in the Andes and the mountains of Central America, from Chile to Mexico.35. Frullania ringens Spruce
On bark of trees, 3400 m. Recorded from Cauca. World distribution: northern Andes (Colombia, Ecuador).
Frullania ringens (subg. Chonanthelia) is recognized by the large, 2-3-toothed laminate portion of the lobule, the long, the decurrent beak of the lobule, the large, rounded underlaves with 2 very large auricles which are usually overlapping each other, and 10-keeled perianths (Yuzawa 1991). Frullania ringens is most closely related to F. ecklonii but differs from the latter by the lobule with a long beak and toothed lamina (beak ± lacking and lamina entire in F. ecklonii) and the larger underleaf auricles, which are often overlapping each other (not overlapping in F. ecklonii).
36. Frullania riojaneirensis (Raddi) Ångstr.
On bark and rock, 500-3800 m. Recorded from Antioquia, Boyacá, Caldas, Casanare, Cauca, Cesar, Chocó, Cundinamarca, Huila, Magdalena, Quindío, Risaralda, Santander and Tolima. World distribution: pantropical.
Frullania riojaneirensis (subg. Chonanthelia) is a very common species of lower montane elevations and is recognized by the large laminate portion of the lobule (to be examined on the leaves of the main stem) with a flat margin and distinct beak, and the 4-keeled perianth.
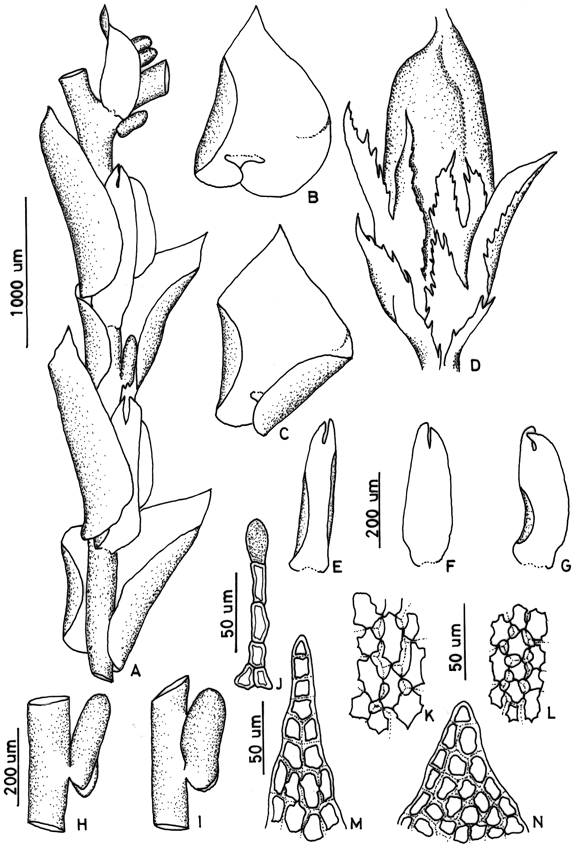
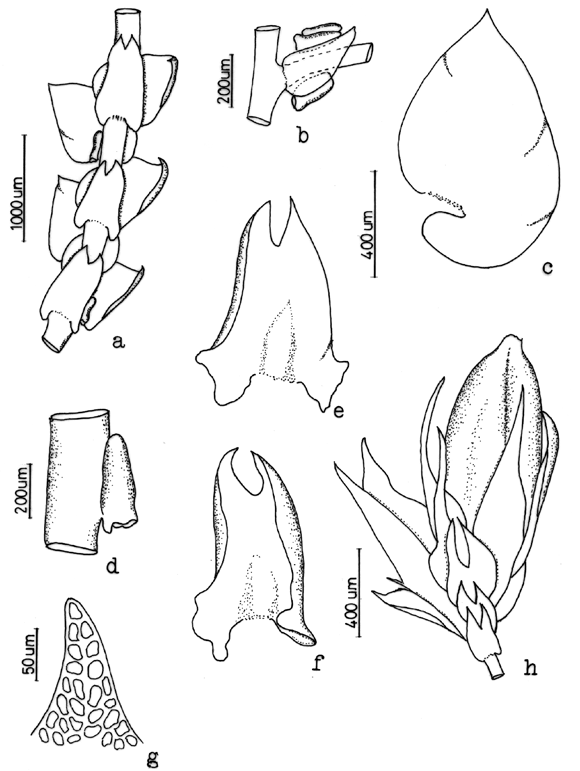
37. Frullania setigera Steph. Fig. 11
]]> On bark of trees, soil, rocks and logs, 900-3600 m. Recorded from Antioquia, Boyacá, Cauca, Chocó, Huila, Santander and Valle. World distribution: scattered in tropical America.
38. Frullania sphaerocephala Spruce
On bark of trees in montane cloud forest areas and páramo, 1700-3900 m. Recorded from Cundinamarca, Meta, Quindío, Risaralda and Tolima. World distribution: tropical Andes from Bolivia to Colombia.
Frullania sphaerocephala (subg. Chonanthelia) resembles F. ecklonii by the pluriplicate perianth, the smooth, entire leaves and underleaves, and the lobule with a large appendage without distinct beak, but is easily separated from F. ecklonii by the large, inflated female bracts and bracteoles, which are closely imbricate and incurved, forming a globose head. Morover, F. ecklonii is monoicous whereas F. sphaerocephala is dioicous. By its peculiar, ball-shaped female involucres F. sphaerocephala cannot be confused with any other neotropical species of Frullania. Further characteristics of F. sphaerocephala are the minute, 1(-2)-celled stylus, the large trigones of the leaf cells and the large auricles of leaves and underleaves. The species is characteristic of upper montane cloud forests and shrubby páramo, above 2500 m.
39. Frullania standaertii Steph.
]]> On bark of trees, 1500-4000 m. Recorded from Cundinamarca and Santander. World distribution: in páramos of the tropical Andes from Colombia to Bolivia; also in East Africa (Kenya).40. Frullania tetraptera Nees & Mont.
On bark of trees, 2000-3800 m. Recorded from Cundinamarca, Meta and Risaralda. World distribution: common at high elevations in Central America and the Andes, from Mexico to Chile and Argentina.
Frullania tetraptera (subg. Chonanthelia) is a characteristic species of the páramo and is recognized by its rather small size (plants ca. 1-1.4 mm wide), the short laminate portion of the lobule, the very small stylus, the narrow underleaves that are hardly wider than the stem, and the monoicous sexuality (Yuzawa, 1991). The species has sometimes been confused with F. cuencensis but the latter species has larger underleaves and is dioicous. Frullania tetraptera is also very similar to F. pluricarinata, which is a poorly known species from high elevation in the Andes, but the latter species has a pluriplicate perianth (see Key).
41. Frullania tunguraguana Clark & Frye
On soil, 1600 m. Recorded from Santander. World distribution: scattered in the tropical Andes (Venezuela to Argentina) and in Mexico.
Frullania tunguraguana (subg. Chonanthelia) is characterized by the pluriplicate perianth and the distincly beaked lobule with a long laminate portion (longer than the sac) whose margins are entire and smooth, not folded or undulate. Also, the margins of leaves and underleaves in this species are never undulate like in the closely related F. arecae. Frullania tunguraguana somewhat resembles F. riojaneirensis but in the latter species the perianths have only 4 keels while those in F. tunguraguana have more 12-14 keels (Yuzawa, 1991). Frullania tunguraguana is known from only very few collections in spite of its wide neotropical distribution.
42. Frullania winteri Steph.
On bark of trees, 1700-2500 m. Recorded from Boyacá, Quindio, Risaralda and Santander. World distribution: northern Andes, Mexico. ]]>
Frullania winteri (subg. Chonanthelia) is recognized by the lobule with a short laminate portion whose lateral margin is folded and projecting upwards towards the sac. Two varieties have been recognized in this species: var. winteri with a relatively large stylus (10-13 cells long, 5-10 cells wide) and var. vanderhammenii (Haarbrink) Yuzawa with a smaller stylus (5-8 cells long, 1-2 cells wide). The material from Colombia and Ecuador belongs to var. vanderhammenii (Yuzawa, 1991). The species is known from only few collections.Doubtful taxa
Frullania armata Herz. - Recorded from Cesar and Magdalena
Frullania atropurpurea Steph. - Recorded from Cundinamarca
Frullania crenulifolia Jack & Steph. - Recorded from Antioquia
Frullania granatensis Gottsche - Recorded from Cundinamarca
Frullania guadalupensis Gottsche ex Steph. - Recorded from Magdalena
Frullania harpantha Herz. - Recorded from Cundinamarca
Frullania moritziana Lindenb. & Gott. - Recorded from Risaralda
Frullania occanniensis Steph. - Recorded from Antioquia ]]>
ACKNOWLEDGMENTS
We thank Dr. Lars Hedenas, Stockholm, for the loan of the type material of Frullania intumescens and Mr. Denis Lamy for the permission to reproduce some figures from Cryptogamie, Bryologie.
LITERATURE CITED
1. BONNER, C. E. B. 1965. Index Hepaticarum V. Delavayella to Geocarpus. Weinheim. 480 pp. [ Links ]
2. GOTTSCHE, C. M. 1864. Hepaticae. In: J. J. Triana, J. J. & J. E. Planchon, Prodromus florae novo- granatensis. Annales des Sciences Naturelles 5, Botanique 1: 95-198. [ Links ]
3. GRADSTEIN, S.R. 2008. Annotated catalogue of liverworts and hornworts of the Galapagos Islands. Bibliotheca Lichenologica 99: 143-162. [ Links ]
4. GRADSTEIN, S. R. & D. P. COSTA. 2003. The Hepaticae and Anthocerotae of Brazil. Memoirs of the New York Botanical Garden 87: 1-317. [ Links ]
5. GRADSTEIN, S. R. & W. H. A. HEKKING. 1979. Studies on Colombian Cryptogams IV. A Catalogue of the Hepaticae of Colombia. Journal of the Hattori Botanical Laboratory 45: 93-144. [ Links ]
6. GRADSTEIN, S.R. & A.-L. ILKIU-BORGES. 2009. Guide to the Plants of Central French Guiana. Part IV. Liverworts and Hornworts. Memoirs of the New York Botanical Garden 76,4: 1-140. [ Links ]
7. GRADSTEIN, S. R. & J. URIBE-M. In press. Marchantiophyta. In: Bernal, R., M. Celis & S. R. Gradstein (eds.), Catalogue of the Plants of Colombia. Bogotá. [ Links ]
8. GRADSTEIN, S. R., S. P. CHURCHILL & N. SALAZAR ALLEN. 2001. Guide to the Bryophytes of Tropical America. Memoirs of the New York Botanical Garden 86: 1-577. [ Links ]
9. HATTORI, S. 1973. Two South American Frullanias, F. mirabilis and F. pendulostyla. Journal of Japanese Botany 48: 104-110. [ Links ]
10. HATTORI, S. 1980. Notes on Asiatic species of the genus Frullania, Hepaticae XII. Journal of the Hattori Botanical Laboratory 47: 85-125. [ Links ]
11. HENTSCHEL, J., M.J. VON KONRAT, T. PÓCS, A. SCHÄFER-VERWIMP, A. J. SHAW, H. SCHNEIDER & J. HEINRICHS. 2009. Molecular insights into the phylogeny and subgeneric classification of Frullania Raddi (Frullaniaceae, Porellales). Molecular Phylogenetics and Evolution 52: 142-156. [ Links ]
12. HERZOG, T. 1942. Beiträge zur Kenntnis neotropischer Bryophyten. Beihefte zum Botanischen Centralblatt 61: 559-590. [ Links ]
13. HERZOG, T. 1955. Hepaticae aus Columbia und Peru. Feddes Repertorium 57: 156-203. [ Links ]
14. HÖRANDL, E. & T. STUESSY. 2010. Paraphyletic groups as natural units of biological classification. Taxon 59:1641-1653. [ Links ]
15. JACK, J. B. & F. STEPHANI. 1892. Hepaticae Wallisianae. Hedwigia 31: 11-27. [ Links ]
16. KESSLER, M. 2000. Elevational gradients in species richness and endemism of selected plant groups in the central Bolivian Andes. Plant Ecology 149: 181-193. [ Links ]
17. KLUGE, J., M. KESSLER & R. DUNN. 2006. What drives elevational patterns of diversity? A test of geometric constraints, climate, and species pool effects for pteridophytes on an elevational gradient in Costa Rica. Global Ecology and Biogeography 15: 358-371. [ Links ]
18. KUNTH, C. S. 1822. Synopsis Plantarum, quas in intere ad plagam aequinoctialem orbis novi collegerunt A. de Humboldt et Am. Bonpland 1: 40-45. Paris. [ Links ]
19. LEÓN-YÁNEZ, S., S. R. GRADSTEIN & C. WEGNER. 2006. Hepáticas y Antoceros del Ecuador. Herbario QCA, Quito. 101 pp. [ Links ]
20. PINZÓN, M., E. L. LINARES & J. URIBE-M. 2003. Hepaticas del Medio Caquetá (Amazonia Colombiana). Caldasia 25: 297-311. [ Links ]
21. ROBINSON, H. 1967a. Preliminary studies on the bryophytes of Colombia. Bryologist 70:1-61. [ Links ]
22. SCHUSTER, R. M. 1992. The Hepaticae and Anthocerotae of North America V. Chicago. 854 pp. [ Links ]
23. SPRUCE, R. 1884. Frullania. En: Hepaticae Amazonicae et Andinae. Trans. Proc. Bot. Soc. Edinburgh 15:3-59. [ Links ]
24. STEPHANI, F. 1901-05. Species Hepaticarum II. Geneva. [ Links ]
25. STOTLER, R. E. 1969. The genus Frullania subgenus Frullania in Latin America. Nova Hedwigia 18: 397-555. [ Links ]
26. URIBE, J. 2004a. Type studies on Frullania subgenus Meteoriopsis (Hepaticae). II. On F. apollinarii and F. paranensis. Caldasia 26: 89-93. [ Links ]
27. URIBE, J. 2004b. Estudios en ejemplares tipo del género Frullania. III. Sobre F. meridana Steph. y F. setigera Steph. Tropical Bryology 25: 9-34. [ Links ]
28. URIBE, J. 2006. Type studies on Frullania subgenus Meteoriopsis (Hepaticae). Frullania dulimensis sp. nov. from Colombia. Cryptogamie, Bryologie 27:309-312. [ Links ]
29. URIBE, J. 2008. Monografia del subgenero Meteoriopsis del género Frullania (Frullaniaceae, Marchantiopsida). Caldasia 30: 49-94. [ Links ]
30. URIBE, J. & S. R. GRADSTEIN. 1998. Catalogue of the Hepaticae and Anthocerotae of Colombia. Bryophytorum Bibliotheca 53: 1-99. [ Links ]
31. URIBE M., J. & S. R. GRADSTEIN. 1999. Estado del conocimiento de la flora de hepáticas de Colombia. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 23, 87:315-318. [ Links ]
32. URIBE, J. & S. R. GRADSTEIN. 2003. Type studies on Frullania subgenus Meteoriopsis. I. Cryptogamie, Bryologie 24: 193-207. [ Links ]
33. VANDENBERGHEN, C. 1976. Frullaniaceae (Hepaticae) africanae. Bulletin du Jardin Botanique National de Belgique 46: 1-220. [ Links ]
34. YUZAWA, Y. 1991. A monograph of subgen. Chonanthelia of gen. Frullania (Hepaticae) of the World. Journal of the Hattori Botanical Laboratory 70: 181-291. [ Links ]
35. YUZAWA, Y. & N. KOIKE. 1989. Studies of the type specimens of Latin American Frullania species (1). Journal of the Hattori Botanical Laboratory 66: 343-358. [ Links ]