
RUBÉN ANTONIO MONTES
JOSÉ SAN-JOSÉ †
GERARDO ANTONIO AYMARD
Universidad Simón Bolívar, Departamento de Estudios Ambientales, Valle de Sartenejas, AP 89000, Caracas, Venezuela. ramontes@usb.ve
Instituto Venezolano de Investigaciones Científicas, Centro de Ecología, Alto de Pipe, AP 20632, Caracas, Venezuela.
]]> Universidad Nacional Experimental de los Llanos Occidentales “Ezequiel Zamora”, UNELLEZ. Herbario Universitario (PORT), Mesa de Cavacas, Portuguesa, Venezuela. gaymard@cantv.netRESUMEN
Se estudiaron las interrelaciones entre la vegetación y el relieve en dos topo-secuencias correspondientes al Parque Nacional Aguaro-Guariquito, estado Guárico, Venezuela. La primera es un paisaje de altiplanicie de Mesa cubierta con una sabana bien drenada (sitio 1). En las aéreas disectadas de esta altiplanicie se presentan geofracturas con suelos permanentemente saturados donde se desarrollan los morichales (sitio 2). La otra topo-secuencia corresponde a una planicie eólica con médanos, cuyos suelos provienen de materiales originados de la altiplanicie de Mesa y están cubiertos por sabanas húmedas (sitio 3), las cuales permanecen anegadas durante tres meses (junio - agosto) con una lámina de agua de 0.25 ± 0.10 m. Las zonas más deprimidas en esta topo-secuencia conducen a condiciones temporales de extremo anegamiento, donde se presenta el congrial (sitio 4), una comunidad dominada por Acosmium nitens y el saladillal (sitio 5), una vegetación caracterizada por la presencia de Caraipa llanorum. El congrial es anegado durante siete meses (junio - diciembre), con una lámina de agua de 1.02 ± 0.10 m. En contraste, el saladillal es inundado por seis meses (junio - noviembre) y la lámina de agua alcanza una altura de 0.80 ± 0.10 m. Las comunidades anegadas (sitios 2-5) se encuentran sobre suelos más ácidos (4.0 4.6 unidades) que los característicos de la sabana bien drenada (sitio 1; 5.0 unidades). El pH estuvo asociado con la concentración de aluminio. El contenido de materia orgánica fue mayor en el sitio anegado (8.59 %) que en los estacionalmente inundables y en el bien drenado (0.7- 1.0 %). El hábitat anegado es restrictivo de la biodiversidad. El congrial, el saladillal, la sabana húmeda y el morichal presentan 52, 52, 56 y 74 especies, respectivamente. En contraste, la sabana bien drenada presenta 114 especies. La similitud entre el congrial y el saladillal fue de 56 %. En relación con las otras comunidades, la similitud fue menor del 38 %. Los resultados indican que las formas de vidas Teroherbae y Teroculmi (e.g., especies anuales de gramínea, ciperácea y herbácea) conforman en promedio un 43 % del espectro de las formas biológicas.
Palabras clave. Sabanas de los llanos centrales de Venezuela, Caraipa llanorum, Acosmium nitens, topo-secuencias.
ABSTRACT
We studied the relationships between vegetation and topography in two topo-sequences corresponding to the National Park Aguaro-Guariquito, Guárico State, Venezuela. The first is a high plain or Mesa landscape covered by a well-drained Savanna (site 1). In the dissected areas of this plateau geofractures occur with permanently saturated soils where the palm Mauritia flexuosa (morichals) grows (site 2). The other topo-sequence corresponds to an eolic plain with soils consisting of materials originating from the Mesa plateau and covered by wet savanna (site 3), which remains waterlogged for three months (June-August) with a water level of 0.25 ± 0.10 m. The most deprived areas in this topo-sequence feature temporary conditions of extreme flooding in which a community dominated by Acosmium nitens (congrial) occurs (site 4), and a vegetation characterized by the presence of Caraipa llanorum (saladillal) (Site 5). The congrial remains flooded for seven months(June-December), with a high water level of 1.02 ± 0.10 m. In contrast, the saladillal remains flooded for six months (June-November) and the water level reaches a height of 0.80 ± 0.10 m. Flooded communities (sites 2-5) are on more acidic soils (4.0 - 4.6 units) than those characteristic of the well-drained savanna (site 1; 5.0 units). The pH was associated with the aluminum concentration. The organic matter content was higher in flooded (8.59%) than in seasonally flooded and well-drained sites (0.7- 1.0 %). The flooded habitat restricts biodiversity. The congrial, saladillal, wet savanna and morichal featured 52, 52, 56 and 74 species, respectively. In contrast, well-drained Savanna has 114 species. The similarity between the saladillal and congrial was 56%. For other communities, similarity was less than 38%. The results indicate that life-forms of Teroculmi and Teroherbae (i.e., annual species of grasses, sedges and herbs) account for 43% of the average of the spectrum of biological forms.
Key words. Central Venezuelan savannas, Caraipa llanorum, Acosmium nitens, topo-sequences.
Recibido: 12/09/2012
Aceptado: 10/09/2013
INTRODUCCIÓN
]]> En América del Sur se encuentran extensas planicies y altiplanicies cubiertas por la ecorregión conocida como los Llanos del Orinoco (Colombia-Venezuela) dominada por una vegetación de sabana, el sistema Río Branco-Rupununi (Guyana-Brasil), las regiones de las cuencas altas de los ríos Manapiare-Ventuari (Venezuela), las sabanas de Guyana, Surinam y la Gran Sabana-Venezuela, sabanas de Moxo en Bolivia y los Cerrados Brasileros (Fanshawe 1952, Heyligers 1963, Blydenstein 1967, Ratter 1980, San José & Montes 1991, Haase & Beck 1989, Montes & San José 1995, Schargel 2007).La cuenca del río Orinoco es la mayor unidad de relieve homogéneo ubicada al norte de América del Sur, con un área de drenaje de 1.1 x 106 km2 y un promedio anual de descarga de 36 x 103 m3 s-1 (Meade et al.. 1983). La cuenca está conformada por una planicie aluvial, cuya alturas oscilan entre 50 y 300 metros sobre el nivel del mar (msnm), originada por sedimentos provenientes de regiones circundantes más altas durante los cambios climáticos ocurridos en el Cuaternario (Schargel, 2007). Desde el punto de vista geoestructural, está región se encuentra conformada por tres provincias tectónicas (Stallard et al. 1990). La primera incluye las altiplanicies aluviales al norte del río Orinoco y sus sedimentos provienen de la Cordillera de la Costa y del Macizo Guayanés (COPLANARH 1974, Malagón 1977). Los sedimentos de la altiplanicie fueron depositados durante el Plio-Pleistoceno, en la Formación Mesa (Hedberg & Pyre 1944). Mientras que, en la segunda y tercera provincias situadas a lo largo de las márgenes sur y suroeste de la cuenca del Orinoco, los sedimentos corresponden a la Cordillera de Los Andes y en menor proporción al Macizo Guayanés. En estas planicies, las interacciones topográficas e hídricas determinan topo-secuencias con la presencia de un grupo de suelos que difieren en propiedades físicas y químicas (Malagón 1977, San José & Montes 1991). En esas formas de relieve se distribuyen un conjunto de comunidades vegetales, con tipo fisionómico y composición florística que se caracterizan por una matriz herbácea dominadas por gramíneas. Las topo-secuencias están caracterizadas por la presencia de: a) una mesa freática que aflora sobre la superficie (e.g., catena edáfica, ver Bushnell 1942, Boul et al. 1989) y b) la acumulación hídrica en las depresiones del paisaje, que prolonga la temporada húmeda. Ambas topo-secuencias son consideradas como determinantes de patrones de distribución espacial de sabanas observado en las planicies de América del Sur (Beard 1953). En los Cerrados centrales de Brasil, el patrón de la vegetación ha sido relacionado con las fluctuaciones temporales del nivel freático (Oliveira-Filho et al. 1990). Así mismo, el tipo de vegetación presente en los llanos del Moxo, Bolivia, ha sido asociado con la interacción del relieve, régimen hídrico y tipo de suelos (Haase & Beck 1989, Haase 1990, 1992). En las sabanas de Suriname, las interacciones entre la humedad del suelo y textura son las principales determinantes de la variación en la composición florística (Van Donselaar 1965). Resultados de los estudios realizados en el ecosistema de las sabanas del Orinoco, señalan que la distribución espacial de la vegetación sobre el relieve responde diferencialmente a la cronología de la deposición de aluviones de los ríos y a la temporalidad del clima (Mazorra et al. 1987, Montes & San José 1995). En las topo-secuencias del Orinoco, donde los suelos permanecen saturados durante todo el año, se observan comunidades dominadas por la palma Mauritia flexuosa (morichales). Mientras que, en topo-secuencias donde ocurren acumulación de las aguas de precipitación y escorrentía superficial, se observa la presencia de comunidades de palmares dominados por Copernicia tectorum ("palma llanera") y sabanas sin elementos leñosos.
El presente estudio se realizó en la margen izquierda del río Orinoco, región donde los paisajes ofrecen una oportunidad única para estudiar el patrón de la vegetación, debido a relieves sometidos a diferentes procesos de modelado, bajo un mismo clima regional. En los llanos del Orinoco, la información acerca de las condiciones hídricas y edáficas que determinan la secuencia vegetacional en los paisajes topo-secuenciales es escasa (Montes et al. 1987, Ramia & Ortiz 2006). El objetivo del presente trabajo, fue evaluar si las variaciones en la composición florística presentes en dos topo-secuencias paisajísticas (e. g., altiplanicie de mesa y planicie eólica) pueden ser atribuidas a las interacciones entre las características fisiográficas, edáficas y niveles de inundación de los paisajes.
MATERIALES Y MÉTODOS
Área del estudio
Este estudio se realizó en el Parque Nacional Aguaro-Guariquito, Estado Guárico, Venezuela. El sistema cubre una zona de 569.000 ha, designadas a la protección y preservación de una amplia gama de ecosistemas llaneros. Estos sistemas merecen especial atención, por sus recursos hidráulicos, refugios de fauna, germoplasmas y asentamientos humanos (Montes et al. 1987). Esta región pertenece a la Provincia Fisiográfica de los Llanos, específicamente a la región natural de los Llanos Centrales (Berroterán 1985). Los paisajes topo-secuenciales del área del estudio están representados por la altiplanicie de Mesa (subregión Llanos Altos Centrales) y planicie eólica (subregión de los Llanos Bajos Centrales). El origen de esta última, es el producto de la deposición de sedimentos provenientes de la erosión hídrica y eólica de la altiplanicie. En esos paisajes, los procesos pedogenéticos, han generado un conjunto de unidades edáficas, entre las cuales se incluyen los órdenes Ultisoles, Oxisoles, Entisoles, Inceptisoles y Molisoles (Malagón 1977, Berroterán 1985).
El clima es megaisotérmico con un promedio anual de precipitación de 1249 mm, y una temporada de crecimiento distribuida entre los meses de abril y noviembre cuando el 98 % de la precipitación ocurre. La temperatura promedio mensual varía desde 29.0 (Abril) hasta 26.5ºC (Diciembre). La evaporación promedio anual de la tina es 2280 mm, lo cual refleja una proporción de 2:1 en comparación con los datos de precipitación.
En la cuenca Aguaro-Guariquito, se seleccionaron dos topo-secuencias de 5 km2 cada una (Figura 1).
Topo-secuencia A
La vegetación predominante, es una sabana bien drenada (sitio 1; 8˚ 25' 49" Norte, 66˚ 30`5" Oeste), donde coexiste una matriz de gramíneas asociadas con elementos leñosos. El componente herbáceo dominante es el Trachypogon spicatus y otros géneros graminosos tales como; Axonopus, Panicum y Paspalum. Los elementos leñosos incluyen Curatella americana, Bowdichia virgilioides y Byrsonima crassifolia. En las áreas disectadas de la altiplanicie de la Mesa ocurren los morichales (sitio 2, 8˚ 25' 36" Norte, 66˚ 30' 33" Oeste) sobre geofracturas, donde los suelos están permanentemente saturados con la mesa freática (e.g., una catena) (González 1987, Mazorra et al. 1987, Montes & San José 1995). Bajo estas condiciones ocurre la palma Mauritia flexuosa y un conjunto de gramíneas, ciperáceas y hierbas.
Topo-secuencia B
Los suelos de esta topo-secuencia se encuentran sobre una Planicie eólica, los cuales, corresponde a acumulaciones de arena en formas de dunas, producto de la influencia de un clima semiárido que ocurrió en el Pleistoceno superior (COPLANARH 1974). La vegetación predominante sobre los médanos y bajíos mejor drenados corresponde a una sabana húmeda con la especie dominante de gramínea del genero de Trachypogon spicatus con una cobertura variable de arboles achaparrados y arbustos (sitio 3, 7˚ 58' 44" Norte, 66˚ 28' 35" Oeste). En el área más deprimida de la topo-secuencia, los suelos limosos y arcillosos interactúan con la pendiente menor de los 5%, generan condiciones de anegamiento durante la temporada de las lluvias. En esas condiciones contrastantes, se pueden observar dos comunidades leñosas contiguas separadas a unos 3 km, una llamada localmente congriales, (sitio 4, 07˚ 58' 6" Norte, 66˚ 29' 12" Oeste) dominada por la especie arbórea Acosmium nitens (Vogel) Jakovlev, y la otra los saladillales (sitio 5, 7˚ 58' 6" Norte, 66˚ 28' 16" Oeste) dominadas por la especies arbórea Caraipa llanorum Cuatrec. (Montes et al. 1987).
Lista de las especies vegetales y el espectro de las formas de vida
Se seleccionó en cada comunidad (sitios 1 - 5) una parcela de 1 ha donde se efectuó un inventario florístico durante cinco años. Se cotejaron los especímenes estériles, con materiales fértiles presentes en las áreas adyacentes. Se depositaron los especímenes en el Herbario Nacional de Venezuela (VEN) y en el de Missouri Botanical Garden, St. Louis, USA (MO).
Recientemente, se han sugerido y realizado numerosos cambios a nivel de familias y géneros. Actualmente, las angiospermas representan el primer grupo de organismos que han sido reclasificado a través de estudios moleculares, y de análisis filogenéticos basados en secuencias de "plastid rbcL" y "atpB genes" (Angiosperm Phylogeny Group 2003, 2009). Por lo tanto, en el presente trabajo, se han empleado las actualizaciones aceptadas por la gran mayoría de los botánicos; por ejemplo: Asclepiadaceae como parte de Apocynaceae, Cochlospermaceae en Bixaceae, los géneros Casearia y Buchnera ubicados en Salicaceae y Orobanchaceae, respectivamente, y Malvaceae unificando a Bombacaceae, Sterculiaceae y Tiliaceae.
Se determinó la similitud florística entre las comunidades vegetales, mediante el cálculo del Índice de Similitud (IS) de Sorensen (1948), IS = 2Sab / (Sa + Sb), donde Sa es el número de especies presentes en la muestra A; Sb el número de especies presentes en la muestra B y Sab el número de especies comunes en ambas muestras.
Se elaboró el espectro de la forma de vida mediante el sistema propuesto por Vareschi (1966). La aplicabilidad del sistema a la vegetación tropical fue comprobada por Montes & San José (1995).
]]> Análisis de suelos a través de las topo-secuenciasEn cada comunidad vegetal, se colectaron 10 muestras compuestas de 500 g de suelo hasta una profundidad de 0.30 m . Las muestras de suelo se desecaron al aire y se tamizaron por una criba de 2 mm de diámetro. Se analizaron, química y físicamente en términos de: pH mediante un electrodo de vidrio en una mezcla de suelo:agua 2:1; porcentaje de carbón orgánico medido por método de Walkley & Black (Jackson 1958); nitrógeno total (N) determinado por el método de micro-kjeldhl (Bremner 1965); fósforo disponible (P) extraído mediante el método de Olsen y su concentración determinada por el método colorimétrico del molibdeno-vanadato (Olsen & Dean 1965); cationes intercambiables (C.I) extraídos con una solución 1N NH4Cl y su concentración determinada por colorimetría de absorción atómica con un Spectrofotometro Varian AA-6.
Altura de la lámina de agua
En cada sitio de estudio, se seleccionaron mensualmente al azar 50 puntos donde se midió la altura de la lámina de agua durante un año.
RESULTADOS
Contenido de nutrientes en los suelos
La composición física y la química de los suelos de las topo-secuencias reflejaron las diferencias del material parental. La fertilidad de los suelos correspondientes a las topo-secuencia A y B fue representada mediante gráficos poligonales propuesto por Alvin & Rosand (1974) (Figura 2).
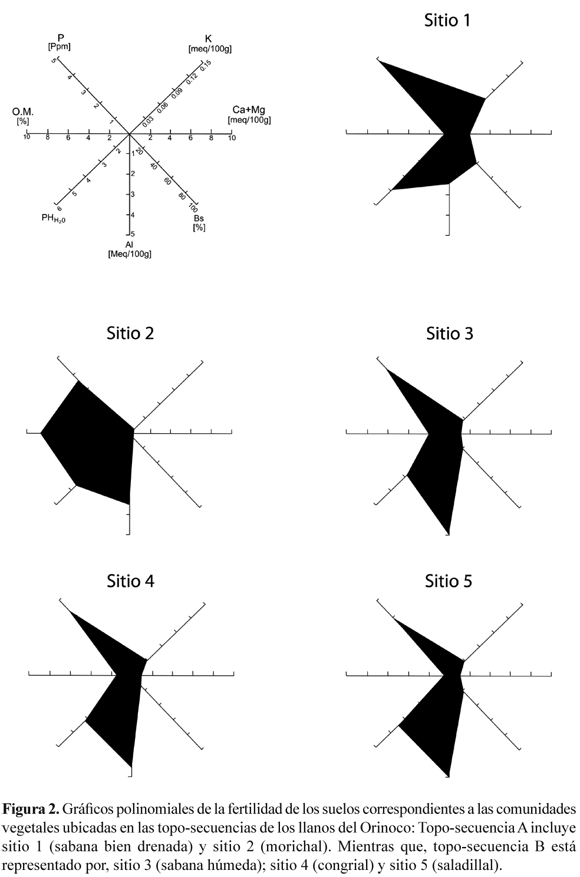
El formato indica que todos los suelos fueron ácidos y pobre en nutrientes. En la topo-secuencia A, ubicada en la altiplanicie de Mesa, los valores de pH disminuyeron desde 5.0, en la sabana bien drenada (sitio 1), hasta 4.0, en el morichal (sitio 2). Mientras que, en la topo-secuencia B, los valores de pH fueron similares, a través de la sabana húmeda (sitio 3; 4.6 ± 0.5), el congrial (sitio 4; 4.3 ± 0.2) y el saladillal (sitio 5; 4.4 ± 0.1). La concentración de aluminio fue mayor en los suelos temporalmente inundados (sitios 3 - 5). Así, las concentraciones de Al en la sabana húmeda, congrial y saladillal, fueron de 4.9 ± 0.9, 4.5 ± 1.0 y 4.9 ± 0.8 meq/100 g de suelo, respectivamente. Mientras que, en la sabana bien drenada y en el morichal fueron de 2.5 ± 0.7 y 3.5 ± 0.9 meq/100g de suelo seco. El fósforo disponible fue muy bajo en todas las comunidades, variando desde 3.3 a 4.8 ppm, con los valores más bajos en los suelos temporalmente inundados, particularmente saladillal. La saturación de las bases fueron muy baja, alcanzando un valor mayor en la sabana bien drenada (48 %), en comparación con los sistemas permanentemente o temporalmente inundados: morichal (9.6 %), sabana húmeda (38.7 %), congrial (19.3 %) y saladillal (25.8 %), respectivamente. El contenido de materia orgánica de los suelos analizados fue mayor en el sistema permanentemente inundado (8.5 %), que en los temporalmente inundables y no inundable (0.7-1.0 %).
]]> Altura y duración de la lámina de aguaEn las comunidades las características de la inundación fueron diferentes (Figura 3).
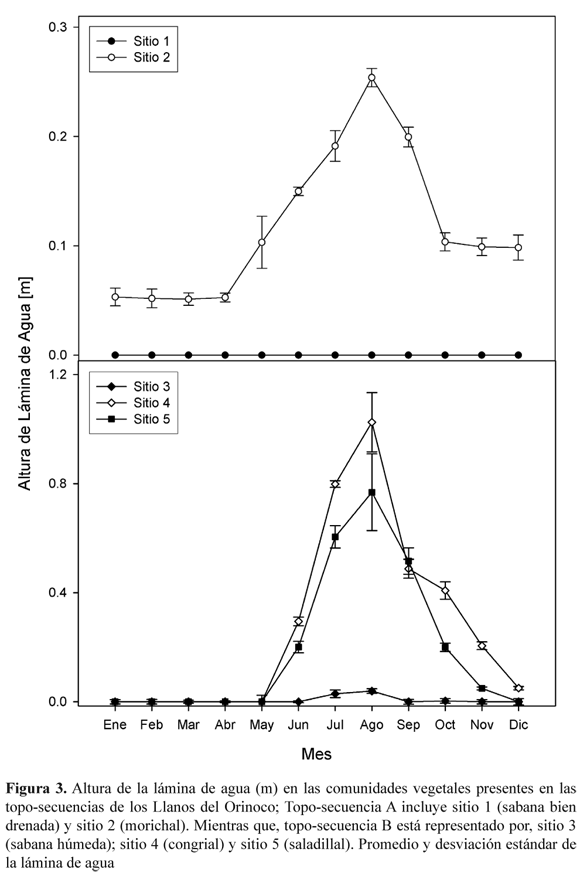
La máxima altura de la lámina de agua ocurrió en Agosto, alcanzando en el congrial (sitio 4) valores de 1.00 ± 0.10 m, seguido por el saladillal (sitio 5) con 0.80 ± 0.10 m y la sabana húmeda (sitio 3) con 0.20 ± 0.10 m, mientras que, en morichal alcanzó 0.31 ± 0.15 m . La duración de la lámina de agua en el saladillal fue de 6 meses (Junio Noviembre), en el congrial 7 meses (Junio Diciembre), y en el morichal permanente.
Composición florística
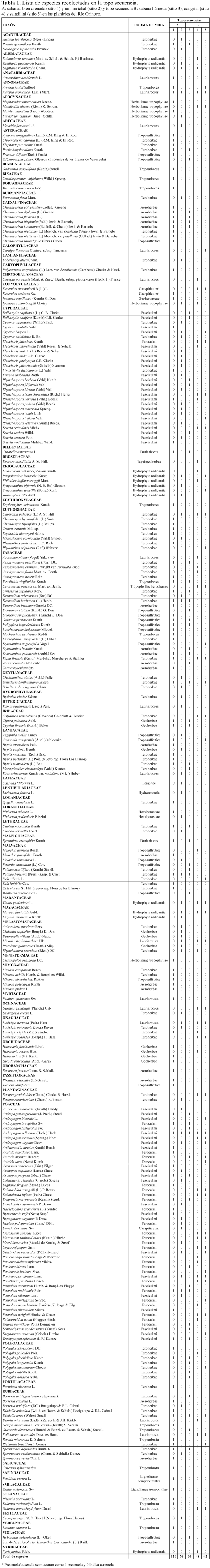
En las dos topo-secuencias (A y B), se identificaron 264 especies pertenecientes a 60 familias (Tabla 1).
Topo-secuencia A
]]> En la topo-secuencia A se encontró la mayor diversidad específica, donde la sabana bien drenada (sitio 1) presentó 114 especies. Las especies dominantes en el estrato herbáceo fueron Trachypogon spicatus (L.F.) Kuntze y Axonopus canescens (Trin.) Pilg., también coexisten árboles aislados y pequeños parches de bosques semidecíduous. Las especies leñosas aisladas más abundantes fueron Curatella americana L., Bowdichia virgilioides Kunth y Byrsonima crassifolia (L.) Kunth. Mientras que, las especies características de los pequeños parches fueron; Cassia moschata Kunth, Copaifera pubiflora Benth., Vochysia venezuelana Stafleu, Zanthoxylum fagara (L.) Sarg. y Godmania aesculifolia (Kunth) Standl. En el estrato arbóreo superior del morichal (sitio 2), caracterizado por la palma Mauritia flexuosa se identificaron 74 especies, también se observo un segundo estrato de elementos arbóreos y arbustos entre 1.5 a 8.0 metros de alto, donde dominan: Ludwigia lithospermifolia (Mich.) Hara, L. nervosa (Poir.), Xylopia aromatica (Lam.) Mart., Cecropia peltata L. y Miconia stephananthera Ule. El estrato herbáceo está dominado por especies propias de áreas con drenaje deficiente, destacan: Cuphea odonellii Lourteig, Desmocellis villosa (Aubl.) Naudin, Rhynchospora velutina (Kunth) Voeck. y Xyris jupicai Rich.Topo-secuencia B
En la topo-secuencia B la sabana húmeda (sitio 3) la domina la gramínea Trachypogon spicatus (L.f.) Kuntze, esta área presentó mayor diversidad específica (56) que el congrial (sitio 4; 52 especies) y el Saldillal (sitio 5; 52 especies). El estrato herbáceo de los congriales y saladillales, es irregular y está representado por Sorghastrum setosum (Griseb.) Hitchc, Panicum aquarum Zuloaga & Murrone y la Ciperácea Eleocharis filiculmis Kunth; el estrato arbóreo está distribuido regularmente. En el congrial, los elementos leñosos están representados por Acosmiun nitens (Congrio) y Couepia paraensis subsp. glaucescens (Hook. F.) Prance y en menor densidad Vochysia venezuelana (Salado), taxón que también se registró en otros congriales localizados al este del área del estudio. El saladillal (sitio 5) está dominado por Caraipa llanorum (Saladillo), son notables las densidades bajas de los arbustos Ouratea guildingii (Planch.) Urb., y Palicourea croceoides Ham. en los congriales y saladillales, taxones propios de suelos sin problemas de drenaje.
Índice de Similitud (IS) entre las comunidades vegetales
Los resultados indican que las comunidades temporalmente inundables de congrial (sitio 4) y saladillal (sitio 5) fueron las más similares (56 %). La sabana bien drenada (sitio 1) y la sabana húmeda (sitio 3) presentaron un 38 % de similitud. Entre los sitios inundables, tales como el morichal (sitio 2) y el congrial (sitio 4), y el morichal y el saladillal (sitio 5), la semejanza fue de 17 y 19 %, respectivamente. Mientras que, entre los sitios 3 y 4; y los sitios 3 y 5, la similitud fue de solo 1 y 6 %, respectivamente. Entre la sabana drenada y el saladillal, las especies comunes fueron Schultesia benthamiana Griseb y Aristida moritzii Henrard (Poaceae), Andropogon selloanus (Hack.) Hack. para todos los sitios. Las diferencias entre los índices reflejaron las variaciones del régimen hídrico.
Espectro de las formas de vida
El espectro de las adaptaciones de las plantas al ambiente de cada comunidad, fue determinado al calcular el número de especies en cada forma de vida en relación al número total de especies (Fig. 4).
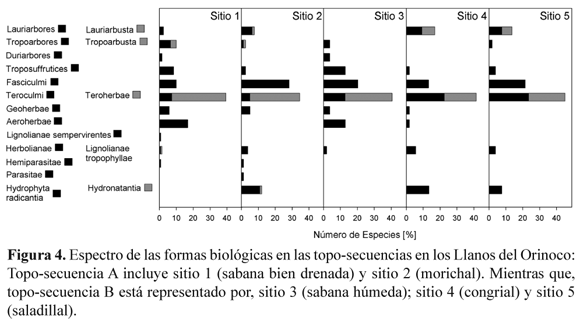
Los Lauriarbores, Lauriarbusta (i.e., plantas de hojas ancha, siempre verde), los Tropoarbores y Tropoarbusta, (i.e., con hojas anchas deciduas), estuvieron presentes en todas las comunidades estudiadas; excepto en la sabana húmeda. Esas formas de vida alcanzaron su máximo valor en el congrial (17 %). Además, la proporción de Troposuffrutices (i.e., pequeños arbustos con dosel senescente durante la temporada de sequía) varió entre 1.8 % en el congrial hasta 13.0 % en la sabana húmeda. Fasciculmi, como las gramíneas y ciperáceas, alcanzó su máxima proporción en el morichal (30 %). Las forma de vida Teroculmi y Teroherbae contemplan especies anuales que incluyen gramíneas, ciperáceas y hierbas. Estas sobrevivieron durante los periodos desfavorables, como semillas y alcanzaron la más alta proporción en el saladillal (44 %) y la menor en la sabana drenada (40 %). Restringida al morichal, Parasitae fue representada por Cassytha filiformis L. En el congrial, se encontró la insectívora Drosera sessiliflora A. St. Hill., y en los sistemas inundables se observó a Hydrophyta radicantia, como especie léntica.
]]> DISCUSIÓNEn este estudio topo-secuencial, se determinó que la interacción entre las diferentes condiciones de relieve, las características edáficas, el grado y duración de la inundación y sequía, parecen haber modificado el balance de hídrico del sistema configurando una amplia gama de tipos fisonómicos y estructuras florísticas. Factores similares han sido reportados en la zona Central de Brasil (Oliveira-Filho et al. 1990). En contraste, los gradientes ambientales en las sabanas bien drenadas están relacionados con las características edáficas y condiciones climáticas (San José & Montes 1991). Estas comunidades (esto es, sabana bien drenada, morichal, sabana húmeda, congrial y saladillal) presentan grupos de especies adaptadas a hábitats y condiciones edáficas extremas. En la altiplanicie de Mesa, la sabana bien drenada (sitio 1), presenta un perfil de suelo estresado hídricamente durante la temporada seca. En condiciones de limitaciones hídricas y baja fertilidad, la tasa de absorción de nutrientes por la vegetación es reducida (Russell 1977). Además, estos suelos pueden presentar resistencia mecánica para el desarrollo radical (San José & Montes 1991). En contraste, los suelos de los morichales (sitio 2) están permanentemente saturados por la mesa freática y presentan un alto contenido de aluminio. Bajo esas condiciones las especies presentan diferentes mecanismos adaptativos a la inundación y alto contenido de aluminio (Mazorra et al. 1987). En la planicie eólica, la vegetación de la sabana húmeda (sitio 3) está asociada a un estrés de saturación hídrica durante tres meses, cuando la lámina de agua alcanza unos 0.25 m . En el congrial, la máxima altura de la lámina de agua fue de 1.02 ± 0.10 m, con una prolongada temporalidad de 7 meses, mayor que en el saladillal, el cual se encuentra a lo largo de las líneas de drenaje. Esta disposición ha sido reportada por Ramia & Ortiz (2006). En un estudio paisajístico de la vegetación, en la región Las Mercedes Cabruta, estado Guárico, describieron resultados similares. Estos autores señalan que en los congriales la densidad arbórea disminuye con la duración de la inundación, la cual se prolonga hasta por 9 meses. Similares resultados han sido reportados para los paisajes de las planicies eólicas limosas, donde los médanos, drenan hacia los morichales y los bosques de galería contiguos a las planicies del río Orinoco (Schargel & Aymard 1993, Schargel 2007, Aymard & González 2007). Los congriales también ocupan las posiciones bajas de las vegas del río Orinoco (esto es, cubetas de decantación), con una prolongada inundación temporal. En contraste, Marin et al. (1998) señalo que en el sur y sureste del estado Guárico las especies de Acosmium nitens, Caraipa llanorum y Duroia micrantha no conforman comunidades monoespecíficas sino que integran bosques temporalmente inundados. En esta localidad, la lámina de agua alcanza una altura mayor ( 1.00 m ) en el mes de Julio y desaparece entre los meses de Diciembre y Enero, con fluctuaciones, marcadas en la periferia. En el sector cóncavo de estos bosques, predominan las plántulas y juveniles de la especie A. nitens, mientras que en la periferia ocurre C. llanorum. Datos suministrados por GBIF (2007) señalan que A. nitens ha sido registrada para las sabanas del Moxo (Bolivia), bosques inundados de Brasil, Guyana y Surinam, donde los niveles de la lámina de agua varía entre 0.30 a 3.0 m de profundidad. Mientras que Blydenstein (1967) señala la presencia de C. llanorum en la llanura aluvial de desborde y morichales de Colombia con inundaciones entre 0.30- 0.40 m de profundidad. En sabanas arboladas-arbustivas al norte del Beni, Bolivia, C. llanorum ha sido reportada sobre planicies inundables y es considerada como constituyente de un grupo de elementos disyuntos con las sabanas del norte del Amazonas (Josse et al. 2007).
El espectro de las formas biológicas descritas en las dos topo-secuencias de los Llanos del Orinoco, reflejó principalmente las adaptaciones de las especies al factor hídrico. En las comunidades sujetas a condiciones de inundación, las formas biológicas Hydrophyta radicantia y Hidronatantia están muy bien representadas. El espectro de las sabanas bien drenadas, contrastó con el de las comunidades temporal y permanentemente inundadas. La duración e intensidad de las condiciones de inundación controlaron la presencia de los elementos leñosos. El exceso hídrico es considerado generalmente restrictivo para el crecimiento de las plantas (Schueler & Holland 2000). Sin embargo, los elementos leñosos característicos del morichal (sitio 2), congrial (sitio 4) y saladillal (sitio 5) presentan adaptaciones fisiológicas y morfológicas a las condiciones de inundación (Parolin 2009) que son convenientes destacar. Fernández et al. (1999) ha reportado que el aparato fotosintético de Acosmium nitens, Symmeria paniculata Benth. y Eschweilera tenuiflora (O. Berg) Miers en las planicies inundables del río Manapire, Venezuela, permaneció integral después de 4 meses de inundación. Las hojas sumergidas se mantuvieron funcionando y contribuyeron con el balance de carbono de las plantas. Herrera et al. (2009) reportó la aclimatización fotosintética en las especies A. nitens, Campsiandra laurifolia Benth, Duroia fusifera Hook. f. y Pouteria orinoensis (Aubrev.) Penn. Marin et al. (1998) midieron los cursos temporales diarios de conductividad estomática en A. nitens, con una media diaria de 1.19 ± 0.38 cm/s, y en C. llanorum de 0.45 ± 0.28 cm/s, los cuales fueron significativamente mayores durante la temporada húmeda. Los autores reportaron valores de potenciales hídricos en A. nitens, los cuales fueron menores en la temporada húmeda y en ningún caso alcanzaron valores < -2.0 MPa. Dezzeo et al. (2003) determinó que en los bosques inundables del río Mapire, Venezuela, la formación de anillos de crecimiento en las especies leñosas (e.g., A. nitens, C. laurifolia, P. orinocensis y Psidium densicomun DC.) ocurre durante los meses de la temporada seca (Noviembre Mayo). Similarmente, Worbes (1997) ha registrado que la producción de biomasa en especies leñosas en las planicies de anegamiento Centrales del Amazonas ocurre durante la fase terrestre. A pesar de las diferencias observadas en el espectro entre las comunidades inundables y no inundables, las especies anuales (Teroculmi y Teroherbae) fueron comunes, al completar su ciclo de vida mediante la latencia de sus semillas. Similares resultados han sido reportados por Linderman (1953) para humedales de Suriname, el río Delaware (EE.UU.) y en los Llanos del Orinoco (Leck & Simpson 1987, Montes & San José 1995).
Los resultados de los análisis florísticos indican que la mayor diversidad se encontró en la sabana bien drenada (sitio 1) con 114 especies seguida por el morichal (sitio 2) con 74 especies. Resultados similares han sido registrados por Montes & San José (1995) para una topo-secuencia equivalente. Mientras que, la sabana húmeda (sitio 3) con 56 especies, el congrial (sitio 4) con 52 especies y el saladillal (sitio 5) con 52 especies, ocuparon los hábitats más restrictivos. La sabana húmeda situada en las proximidades de los congriales y saladillales presentó un número menor de especies, que en las sabanas de Trachypogon spicatus temporalmente inundables en una planicie aluvial (122 especies) (Montes & San José 1995). En relación a la similitud entre las comunidades, los congriales y saladillales fueron las comunidades más semejantes (56 %). Mientras que, la similitud entre la sabana bien drenada y las temporalmente inundables, (i.e., saladillal y congrial), fueron entre 6 y 10 %. Las especies comunes entre la sabana bien drenada y el saladillal fueron Schultesia benthamiana y Aristida moritzii. Las diferencias en el Índice de Similitud reflejan variaciones del régimen hídrico.
Una comparación de la composición florística entre las comunidades presentes en la dos topo-secuencias estudiadas, las reportadas para los llanos del Orinoco y otras localizadas en América del Sur (Montes & San José 1995) indican que el número de familias de Angiospermas fue similar. Las especies Xylopia aromatica y Andropogon selloanus, están presentes en cuatro de las cinco comunidades y representan las especies "generalistas", por su adaptación a los hábitats extremos. En contraste, Caperonia palustris (L.) A St. Hill, Thalia geniculata L., Mayaca fluviatilis Aubl., Miconia stephananthera, Ludwigia nervosa y Panicum parvifolium Lam. constituyen un grupo de especies "especialistas" (Gentry 1986), en virtud que solamente se encuentran en las comunidades de los sitios 2 (morichal), 4 (congrial) y 5 (saladillal) respectivamente. Finalmente, Poaceae y Cyperaceae fueron características de la sabana bien drenada (sitio 1, 12 y 4 %), el morichal (sitio 2, 19 y 20 %), la sabana húmeda (sitio 3, 23 y 9 %), el congrial (sitio 4, 27 y 10 %) y el saladillal (sitio 5, con 35 y 10 %). Las familias Poaceae y Cyperaceae están muy bien representadas en los sistemas inundables, bien drenados de las sabanas del Moxo, en Bolivia (Haase 1990), Humaita, Amazonas (Gottberger & Morawetz 1986) y la gran mayoría de las sabanas de los Llanos del Orinoco (Huber, 2007).
La vegetación de la región estudiada se encuentra sobre suelos oligotrófos, susceptibles a una rápida degradación si son utilizados intensamente. Por lo tanto, es necesario conservar los horizontes superficiales, ya que son la fuente de nutrimentos a la vegetación nativa, resultados similares han sido reportados para la planicie eólica limosa del Cinaruco-Capanaparo, estado Apure, Venezuela (Schargel & Aymard, 1993). La limitada fertilidad natural de los suelos, que combinada con las características topográficas, clima y drenaje restringen sus posibilidades de uso. Hasta la fecha la ganadería extensiva ha sido permitida y necesario un largo periodo de recuperación de la vegetación nativa.
AGRADECIMIENTOS
Este trabajo fue realizado como parte del Proyecto de Bio-Productividad de Sabanas (UNESCO/MAB) del IVIC. Se reconoce el apoyo financiero del Instituto Nacional de Parques y el Decanato de Investigaciones de la Universidad Simón Bolívar. Deseamos darle las gracias por su valiosa colaboración a los Drs. Julian Steyermark y Gerrit Dadvise por la identificación de los especímenes recolectados, Dr. Dirk Thielen (IVIC) de UNISIG por el procesamiento de la imagen SPOT5 y al Sr. Emilio Rodríguez, Guardaparque por su ayuda en las mediciones y colección de muestra de campo. Deseamos también agradecer el apoyo profesional de Nathaly Matute (Centro de Ecología, IVIC).
LITERATURA CITADA
1. Alvin, P. & F. Rosand. 1974. Un novo sistema do representação gráfica da fertilidade dos solos para cacao. Cacau Atualidades 11:2-6. [ Links ]
2. Angiosperm Phylogeny Group 2003. An update of the Angiosperm Phylogeny group classification for the orders and families of flowering plants: APG II. Botanical Journal of the Linnean Society 141: 399-436 [ Links ]
3. Angiosperm Phylogeny Group 2009. An update of the Angiosperm Phylogeny group classification for the orders and families of flowering plants: APG III. Botanical Journal of the Linnean Society 161: 105-121. [ Links ]
4. Aymard, G. & V. González. 2007. Consideraciones generales sobre la composición florística y diversidad de los bosques de los Llanos de Venezuela. En: R. Duno de Stefano, G. Aymard & O. Huber (eds). Catalogo anotado e ilustrado de Flora vascular de los Llanos de Venezuela: 59-72. FUDENA, FUNDACIÓN POLAR, FIBV, Caracas. [ Links ]
5. Beard, J.S. 1953. The savanna vegetation of northern tropical America. Ecological Monographs 23:149-215. [ Links ]
6. Berroterán, J.L. 1985. Geomorfología de un área de Llanos bajos centrales Venezolanos. Boletín Sociedad Venezolana Ciencias Naturales 48(143):31-78. [ Links ]
]]>7. Blydenstein, J. 1967. Tropical savanna vegetation of the Llanos of Colombia. Ecology 48:1-15. [ Links ]
8. Bremner, J.M. 1965. Total nitrogen. In: Black, C (ed.). Methods of soil analysis. American Society of Agronomy. Wisconsin. 1149-1178. [ Links ]
9. Buol, S.W., F.D. Hole & R. McCracken. 1989. Soil genesis and classification. 3rd ed. Iowa State University Press, Ames. 446 p. [ Links ]
10. Bushnell, T.H. 1942. Some aspects of the soil catena concepts. Soil Science Society of America Proceedings 7:466-476. [ Links ]
11. COPLANARH. 1974. Inventario Nacional de Tierras. Regiones Centro-Oriental y Oriental. Publicación No. 35. MARNR. Caracas, Venezuela. [ Links ]
]]>12. Dezzeo, N., M. Worbes, I. Ishii & R. Herrera. 2003. Annual tree rings revealed by radiocarbon dating in seasonally flooded forest of the Mapire river, a tributary of the lower Orinoco river. Plant Ecology 168:165-275. [ Links ]
13. Fanshawe, D.B.1952. The vegetation of British Guiana. A preliminar review. Imperial Forestry Institute, Oxford University. Paper No. 29. [ Links ]
14. Fernandez, M.D., A. Pieters, C. Donoso, C. Herrera, W. Tezara, E. Rengifo & A. Herrera. 1999. Seasonal changes in photosynthesis of trees in the flooded forest of Mapire River. Tree Physiology 19:79-85. [ Links ]
15. Gentry, A. 1986. Endemism in tropical versus temperate plant communities. En: M.E. Soulé (ed). Conservation Biology: 153-181. Sinauer Association, Sunderland, Massachusetts. [ Links ]
16. Global Biodiversity Information Facility (GBIF) 2007. Map of Acosnium nitens. http://www.discoverlife.org/MP/20M?Kind=Acosnium+nitens [ Links ]
17. González, V. B. 1987. Los morichales de los llanos Orientales. Un enfoque ecológico. Cuadernos Corpoven. Caracas. Venezuela. [ Links ]
18. Gottberger, G. & W. Morawetz. 1986. Floristic, structural and phytogeographical analysis of the savannas of Huimata (Amazonas). Flora 178:41-71. [ Links ]
19. Haase, R. 1990. Plant communities of a savanna in northern Bolivia. II. Palm swamps, dry grasslands, and shrubland. Phytocoenologia 18:343-370. [ Links ]
20. Haase, R. 1992. Physical and chemical properties of savanna soil in northern Bolivia. Catena 19:119-134. [ Links ]
21. Haase, R. & S.G. Beck. 1989. Structure and composition of savanna vegetation in northern Bolivia. A preliminary report. Brittonia 41:80-100. [ Links ]
22. Hedberg, H. & A. Pyre. 1944. Stratigraphy of northeastern Venezuela. American Association of Petroleum Geologists Bulletin 28:1-28. [ Links ]
23. Herrera, A., M. Escala & E. Rengifo. 2009. Leaf anatomy changes related to physiological adaptations to flooding in Amazonian tree species. Brazilian Journal of Plant Physiology 21:301-308. [ Links ]
24. Heyligers, P.C. 1963. Vegetation and soil of a white sand savanna in Suriname. The vegetation of Suriname 3:1-148. [ Links ]
25. Huber, O. 2007. Sabanas de los Llanos de Venezuela. En: R. Duno de Stefano, G. Aymard & O. Huber (eds). Catálogo ilustrado y anotado de las plantas vasculares de los Llanos de Venezuela: 73-86. FUDENA, FUNDACIÓN POLAR, FIBV, Caracas. [ Links ]
26. Jackson, M.L. 1958. Soil chemical analysis. Prentice Hall. New York. 498 pp. [ Links ]
27. Josse, G., G. Navarro, F. Encarnación, A. Tovar, P. Comer, W. Ferreira, F. Rodríguez, J. Saito, J. Sanjurjo, J. Dyson, E. Robien de Celis, R. Zarate, J. Chang, M. Ahuite, C. Vargas, F. Paredes, W. Castro, J. Maco & F. Reategui. 2007. Sistemas Ecológicos de la Cuenca Amazónica de Perú y Bolivia. Clasificación y mapeo. NatureServe, Arlington. 95pp. [ Links ]
28. Leck, M.A. & R.L. Simpson. 1987. Seed bank of a freshwater tidal wetland: Turnover and relationships to vegetation change. American Journal of Botany 74:360-370. [ Links ]
29. Linderman, J.C. 1953. The vegetation of the coastal region of Suriname. The vegetation of Suriname 1:1-135. [ Links ]
30. Malagón, D. 1977. Modelos y caracterización de la génesis de los suelos en dos ecosistemas tropicales (Sabana y Selva Pluvial) de Colombia. CIDIAT, Mérida, Venezuela. 84 pp. [ Links ]
31. Marín, Ch. D., L. Hernández & G. De Martino. 1998. Ecología de los congriales de la Estación Experimental Nicolasito (Santa Rita, estado Guárico, Venezuela). I. Medio físico, estructura de la comunidad y microclima en épocas contrastantes. Memorias Sociedad Ciencias Naturales Salle 58:3-18. [ Links ]
32. Mazorra, M.A., J.J. San José, R.A. Montes, J. Garcia-Miragaya & H, Haridasan. 1987. Aluminium concentration in the biomass of native species of the Morichals (swamp palm community) at the Orinoco Llanos, Venezuela. Plant and Soil 102:275-277. [ Links ]
33. Meade, R.H., C.P.Jr. Nordin, D. Perez-Hernandez, B. Mejia & J.M. Perez-Gody. 1983. Sediment and water discharge in río Orinoco, Venezuela and Colombia. En: Proceedings of the second International Symposium on River Sedimentation. Water Resources and Electric Power Press. pp. 1134-1144, Beijing. [ Links ]
34. Montes, R. & J.J. San José. 1995. Vegetation and soil analysis of topo-sequences in the Orinoco Llanos. Flora 190:1-33. [ Links ]
35. Montes, R.A., M. Sebastiani, F. Delascio, J. Arismendi & I. Mesa. 1987. Paisajes-vegetación e hidrografía del Parque Nacional Aguaro-Guariquito, Estado Guárico. Boletín de la Sociedad Venezolana de Ciencias Naturales 41:73-112. [ Links ]
36. Oliveira-Filho, A.T., & F.R. Martins, J. Ratter, & G.J. Shepherd. 1990. Floristic composition and community structure of a central Brazil gallery forest. Flora 184:103-117. [ Links ]
37. Olsen, S.R. & L.A. Dean. 1965. Phosphorous. En: C. Black (ed). Methods of soil analysis: 1035-1048. American Society of Agronomy, Wisconsin. [ Links ]
38. Parolin, P. 2009. Submerged in darkness: adaptations to prolonged submergence by woody species of the Amazon floodplains. Annals of Botany 103:359-376. [ Links ]
39. Ramia, M & R. Ortiz. 2006. Relaciones ecológicas en paisajes de sabana en la región Las Mercedes Cabruta, Estado Guárico. Boletín de la Sociedad Venezolana de Ciencias Naturales 154:62-136. [ Links ]
40. Ratter, J.A. 1980. Notes on the vegetation of fazenda agua limpa ( Brasilia, D.F. Brasil). Royal Botanical Garden, Edinburgh. 115 pp. [ Links ]
41. Russell, R.S. 1977. Plant root systems: Their function and interaction with the soil. McGrow-Hill, London. 258 pp. [ Links ]
42. San José, J.J. & R.A. Montes. 1991. Regional interpretation of environmental gradients which influence Trachypogon savannas of the Orinoco Llanos, Venezuela. Vegetatio 95:21-32. [ Links ]
43. Schargel, R. 2007. Geomorfología y Suelos. En: R. Duno de Stefano, G. Aymard & O. Huber (eds).. Catalogo anotado e ilustrado de la flora vascular de los Llanos de Venezuela: 21-42. FUDENA, FUNDACIÓN POLAR, FIBV, Caracas. [ Links ]
44. Schargel, R. y G. Aymard. 1993. Observaciones sobre suelos y vegetación en la llanura eólica limosa situada entre los ríos Capanaparo y Riecito. Estado Apure. Venezuela. Biollania 9: 119-147. [ Links ]
45. Schueler, T.R. & H.K. Holland. 2000. Practice of watershed protection. Wetter is not always better: flood tolerance of woody species. Center for Watershed Protection, Ellicott City, Maryland. [ Links ]
46. Sorensen, T. 1948. A method of establishing groups of equal amplitude in plant society based on similarity of species content. K. Danske Vidensk. Selsk. 5:1-34. [ Links ]
47. Stallard, R.F., L. Koehnken, & M.J. Johnsson. 1990. Weathering processes and the composition of inorganic material transported through the Orinoco River system. Venezuela and Colombia. The Orinoco River as an ecosystem. En: Convención Anual de la Asociación Venezolana para el Avance de la Ciencia. pp. 81-119, Valencia, Venezuela. [ Links ]
48. Van Donselaar, J. 1965. An ecological and phytogeographic study of northern Surinan savannas. Wentia 14:1-163. [ Links ]
49. Vareschi, V. 1966. Sobre las formas biológicas de la vegetación tropical. Boletín de la Sociedad Venezolana de Ciencias Naturales 26: 504-518. [ Links ]
50. Worbes, M. 1997. The forest ecosystem of the floodplains. En: W.J. Junk (ed.). The Central Amazon Floodplains Ecology of a Pulsing System: Ecological Studies 126, 223-266. Springer, Berlin, Heidelberg, New York. [ Links ]
]]>