Equine herpesvirus 1 and 4 DNA detection in peripheral blood mononuclear cells and trigeminal ganglion of equines: Infection, latency and approximation to neuropathogenesis of the strain
Detecção de DNA tipos de herpesvírus eqüino 1 e 4 nas células mononucleares do sangue periférico e os gânglios trigémeo de cavalos. Infecção, latência e de uma abordagem para o neuropatogénesis da cepa circulante
Julián Ruíz Sáenz1*, MV, MS; Yenny Góez1, MV; Albeiro López Herrera2, Zoot, MV, MS, Dr. Sci.
1Corporación Ciencias Básicas Biomédicas, Universidad de Antioquia, Medellín, Colombia.
2Grupo BIOGEM, Universidad Nacional de Colombia, Sede Medellín. Medellín, Colombia.
(Recibido: 10 septiembre, 2007; aceptado: 6 agosto, 2008)
]]>
Resumen
La infección primaria por Herpesvirus Equino tipos 1 y 4 (HVE-1 y HVE-4) se inicia en el tracto respiratorio superior; luego se produce una viremia primaria en la que intervienen linfocitos B y T, la cual le permite al virus alcanzar otros sistemas orgánicos y producir abortos en el último tercio de la gestación, muerte neonatal de potros y síndromes neurológicos, principalmente por causa del HVE-1. Debido al hallazgo de anticuerpos contra HVE-1 y HVE-4 en caballos de dos departamentos de Colombia, el propósito de este estudio fue detectar la presencia del genoma viral en Mononucleares de Sangre Periférica (MNSP) de caballos seropositivos para HVE-1 y HVE-4 y en ganglios trigéminos de equinos en una planta de faenado del departamento de Antioquia. Por medio de una PCR semianidada se amplificaron los genes que codifican por las glicoproteína Hs (gH) de HVE-1 y B (gB) de HVE-4. El 28 y el 19% de los MNSP contenían el gen de la gH y de la gB, respectivamente. En el 57.8 y 47.7% de los ganglios trigéminos evaluados se logró amplificar los genes gH y gB, respectivamente. Para determinar si la cepa de HVE-1 circulante en el departamento de Antioquia poseía potencial neuropatogénico, se amplificó y secuenció el gen de la DNA polimerasa viral, que puede presentar una mutación asociada con neuropatogénesis. Sin embargo, ninguna de las cepas secuenciadas poseía dicha mutación. Los resultados confirman la presencia de la infección por HVE -1 y HVE-4 en el departamento de Antioquia, lo que sugiere que existen animales con infección latente que podrían ser una fuente de infección para otros animales susceptibles.
Palabras clave: infección latente, patogénesis por herpesvirus, secuenciación de polimerasa viral.
Summary
The infection with Equine Herpesvirus types 1 and 4 (EHV-1 and EHV-4) occurs at the upper respiratory tract. Soon after this takes place a primary cell associated viremia to peripheral blood mononuclear cells (PBMC, mainly on B and T lymphocytes), which allows the virus to reach other organic systems and production of abortions in the last third of gestation, neonatal foal death and neurological syndromes. After primary infection the animals remain latently infected for all the life. Because the presence of antibodies for EHV-1 and EHV-4 in plasma and serum of horses of two departments of Colombia was demonstrated, the objective of the present study as to demonstrate the presence of the viral genome in PBMC from horses diagnosed seropositive for EHV-1 and EHV-4, and in trigeminal ganglion of equines from a slaughterhouse of the Department of Antioquia. By means of a semi-nested PCR, the gene codifying for glycoprotein H (gH) of EHV-1 and gB of EHV-4 were amplified. In PBMC 28 and 19% of gH and of gB amplification were found, respectively; whereas in trigeminal ganglion 57.8 and 47.7% were amplified for gH and gB, respectively. With the aim of assessing whether the circulating strain in the department of Antioquia had a neuropathogenic potential, we amplified and sent to sequencing the gene that encodes the viral DNA polymerase, which could has a mutation that has been associated with neuropathogenic potential. We found that the circulating viral strain in Antioquia does not have such a mutation. The set of our results confirms that infection by EHV is present in the State of Antioquia, Colombia, and that there are equines latently infected which can be a source of infection for other susceptible horses.
Key words: herpesvirus pathogenesis, latent infection, viral polymerase sequencing
Resumo
]]> A infecção primária por Herpesvirus Eqüino tipo 1 e 4 (HVE-1 e HVE-4) inicia no tracto respiratório superior; depois, há uma viremia primária envolvidos no B e linfócitos T, que permite que o vírus chegue a outros sistemas e produtos biológicos abortos no último terço da gestação, morte neonatal de potros e síndromes neurológicas, devido sobretudo ao facto de HVE-1. Devido à descoberta de anticorpos contra o HVE-1 e HVE-4 cavalos em dois departamentos da Colômbia, O objetivo deste estudo foi o de detectar a presença do genoma viral em MNSP cavalo seropositivos para HVE-1 e HVE-4 4 os gânglios do trigémeo de cavalos em um curativo na planta do departamento de Antioquia. Através de uma PCR semianidada foram amplificados genes codificantes para a glicoproteína Hs (GH) HVE-1 e B (GB) de HVE-4. Em 28 e 19% do MNSP contendo o gene para a GH e no Reino Unido, respectivamente. No 57,8 e 47,7% dos gânglios trigêmeo avaliados foi atingido amplificar genes GH e GB, respectivamente. Para determinar se a estirpe do HVE-1 circulantes no departamento de Antioquia tinha potencial neuropatogénico, foi amplificado e sequenciado o gene da DNA polimerase viral, que pode apresentar uma mutação associada com neuropatogénesis. No entanto, nenhuma das cepas seqüenciadas possuía essa mutação. Os resultados confirmam a presença de infecção e de HVE -1 HVE-4, no departamento de Antioquia, sugerindo que há animais com infecção latente que poderia ser uma fonte de infecção para outros animais sensíveis.Palavras chave: infecção latente, patogênese por herpesvírus, seqüenciamento de polimerase viral
Introducción
Los herpesvirus equinos 1 y 4 (HVE-1 y HVE-4), que pertenecen a la subfamilia Alfaherpesvirinae son patógenos altamente prevalentes en diversas regiones del mundo y causan graves pérdidas económicas debidas a un amplio rango de síntomas clínicos que van desde la presentación de una enfermedad respiratoria hasta la inducción de abortos en el último tercio de la gestación, muerte neonatal de potros, ocasionalmente lesiones oculares y daño neurológico, resultando en parálisis del tren posterior (15).
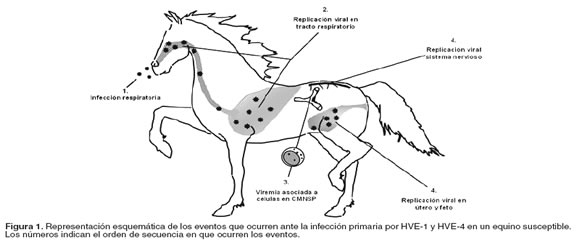
HVE-1 y HVE-4 causan infección sistémica en caballos. La infección primaria se presenta en las células epiteliales del tracto respiratorio superior (19); luego, el virus penetra la barrera epitelial para replicarse en los tejidos profundos del trato respiratorio y nódulos linfoides locales (20); 4 a 6 días después de la primoinfección el virus infecta células mononucleares de sangre periférica (MNSP) resultando una viremia asociada a células, la cual dura 14 días en promedio (15, 42); aunque existen registros de viremias de hasta 27 días en equinos después de la primoinfección (21). Así, los MNSP infectados se convierten en un medio de dispersión del virus en el hospedero, que le permite alcanzar otros sistemas orgánicos, tales como el útero y el sistema nervioso central (véase Figura 1), causando abortos y enfermedades neurológicas. Se ha descrito que tanto la infección del sistema nervioso, como la infección de tracto reproductivo de yeguas, son debidas a la transferencia de virus infecciosos desde los MNSP, a las células endoteliales de los vasos sanguíneos del endometrio y del sistema nervioso (11, 36).
]]> Dado que los MNSP son las células portadoras del HVE en la sangre, éstas juegan un papel crucial en la patogénesis de la infección por éstos virus. La infección in vivo de MNSP fue descrita por primera vez en 1983 por Scott et a, (33). Los autores infectaron ponis y encontraron que el virus permanecía entre 2 y 14 días asociado con los linfocitos T las principales células MNSP infectadas. Cuando los MNSP de ponis infectados fueron cultivados in vitro, en presencia de un agente mitógeno, se incrementó en 10 veces la producción de virus, lo que sugería que el mitógeno promovía la replicación viral (10, 33). Estudios in vitro realizados con monocitos y linfocitos equinos estimulados con diferentes agentes mitógenos e infectados con HVE-1, permitieron demostrar que los linfocitos T CD5+ CD8+ (citotóxicos), son la principal subpoblación celular susceptible a la infección, seguidos por linfocitos B y monocitos (43, 44). Además, se ha sugerido que un estado específico de la fase S y G2/M del ciclo celular, facilita la replicación viral e induce la formación de uniones intercelulares entre las células MNSP y favorece la transmisión del HVE-1 de una célula infectada a las células adyacentes (45).
Luego de la infección primaria, los HVE-1 y HVE-4 establecen una infección latente y permanecen en el hospedero de forma indefinida. En animales de plantas de faenado es posible encontrar infección latente en ganglios trigéminos en aproximadamente el 60 al 80% de la población de caballos, después de una primoinfección (12, 35), por lo cual se ha postulado que los individuos con infección latente son el principal reservorio de la infección para otros caballos susceptibles (1). Durante la infección latente el genoma viral se encuentra en un estado de transcripción restringida, cuando sólo se producen los transcriptos asociados a latencia (LATs, del inglés Latency-Associated Transcripts), sin encontrar proteínas virales y mucho menos partículas virales infecciosas (31). Sin embargo, estudios recientes demostraron mediante el uso de PCR en tiempo real, que la presencia de LATs de HVE-4 en MNSP durante la primera semana posinfección no es un indicativo de establecimiento del estado de latencia en esas células (28).
El genoma de HVE-1 y HVE-4 en estado latente se ha encontrado principalmente en el ganglio trigémino (5, 7, 34), el tejido linfoide que drena el tracto respiratorio y en leucocitos de sangre periférica (4, 8, 35). La infección latente del HVE-1 en leucocitos de sangre periférica se establece predominantemente en linfocitos T citotóxicos (CD5+/CD8+) (35). Por el contrario, estudios con HVE-4 demuestran que este no establece latencia en linfocitos de sangre periférica (6, 28, 48).
Aunque se han descrito diferentes cepas de HVE-1 con mayor o menor asociación con la presentación de abortos y mieloencefalitis (24, 27, 38), las diferencias en el potencial patogénico de las cepas, se correlacionan con diferencias en la capacidad para diseminarse y establecer infección en el endotelio vascular. Al parecer, una mutación puntual presente en la enzima DNA polimerasa del HVE-1 (ORF30), tiene una fuerte asociación con brotes de enfermedad neurológica versus brotes de enfermedad no neurológica in vivo (26); de ahí que mediante estudios de mutagénesis viral se pudo concluir que dicha mutación está directamente implicada en la neuropatogénesis de la enfermedad (16). Por lo tanto, esta mutación se está utilizando como un marcador genético para identificar y predecir el potencial neuropatogénico de las cepas circulantes de HVE-1. En paralelo, se demostró mediante la técnica de PCR en tiempo real, que la magnitud y la duración de la viremia asociada a las células, era mayor en potros infectados con cepas neuropatogénicas versus las cepas no neuropatogénicas (3); también, estudios recientes in vitro confirmaron que las cepas que portan la mutación tiene mayor tropismo por MNSP, principalmente células CD4+ (16), lo que sugiere que la infección de MNSP juega un papel de gran importancia en la neuropatogénesis de la infección.
En un estudio reciente, trabajando con cepas aisladas de diferentes regiones del mundo durante aproximadamente 30 años, y mediante PCR y análisis de secuencia, se demostró que el ORF68 del HVE-1 es particularmente polimórfico y que este puede ser adoptado como un marcador único, debido a que permite distinguir y categorizar los diferentes aislamientos virales de HVE-1 en seis grupos comunes, los cuales presentan gran restricción geográfica (26). Debido a esto, es posible postular una asociación entre cepas de diferentes zonas geográficas y posibles orígenes de introducción o reintroducción de la infección.
En un estudio previo, nuestro grupo de investigación reportó una seropositividad para de 18.8 y 98.7% para HVE-1 y HVE-4, respectivamente, en el departamento de Antioquia (Colombia) (32). Teniendo en cuenta que los MNSP y los ganglios trigéminos de los equinos infectados por el HVE-1 y HVE-4, son las principales células en las cuales se mantiene la infección latente y específicamente son los MNSP las células a partir de las cuales se realiza la transferencia de virus infecciosos a otros sistemas orgánicos, el propósito del presente estudio fue: evaluar la presencia del genoma viral de HVE-1 y HVE-4 en MNSP de equinos previamente diagnosticados como seropositivos, por medio de una PCR semianidada (aceptada por la OIE como prueba diagnóstica para los HVE); determinar la presencia de genoma viral en ganglios trigéminos de equinos de una planta de faenado, lo cual permitiría postular la presencia de infección latente en el departamento de Antioquia (reservorios de la infección viral); y amplificar y secuenciar el ORF30 del HVE-1, con el propósito de determinar el potencial neuropatogénico de la cepa circulante en la región objeto de estudio.
Los resultados confirman por métodos moleculares la presencia de la infección por HVE-1 y HVE-4 en la población equina del departamento de Antioquia; además muestran que es posible detectar genoma viral tanto en MNSP como en ganglio trigémino, por lo que se postula que existen animales con infección latente, los cuales pueden ser fuente de infección para otros equinos susceptibles. Finalmente, por los resultados de secuenciación del ORF30 del HVE-1, se postula que la cepa circulante en Antioquia no posee potencial neuropatogénico.
Materiales y métodos
Aval del Comité de Ética
]]> Esta investigación contó con la autorización del Comité de ética para experimentación con animales de la Universidad de Antioquia (Acta 15 de septiembre 16 de 2004).Colección de muestras
Células mononucleares de sangre periférica (MNSP). La detección de genoma viral en MNSP se realizó en muestras tomadas de los caballos que previamente se habían evaluado para determinar la seropositividad para el HVE-1 y el HVE-4, mediante una prueba de ELISA; en dicho estudio se encontró que de los 96 caballos evaluados en el departamento de Antioquia, 95 fueron positivos para HVE-4 y 18 para HVE-1 (32). Los caballos fueron sangrados por punción en la vena yugular y la muestra fue colectada en tubos de 10 ml con heparina como anticoagulante (Vacutainer Systems® Becton Dickinson, USA). Luego de ser transportadas al laboratorio, las muestras fueron centrifugadas a 640 x g/ 15 min/4 °C, se extrajo el plasma y la capa de leucocitos fue separada y mezclada dos veces con 5 ml de tampón de lisis de eritrocitos (0.85% NH4Cl, 17mM, Tris, pH 7.4) por 5 minutos y centrifugada a 640 x g/5 min (14). El sobrenadante fue descartado y el botón resultante de MNSP fue resuspendido en 0.9% NaCl y congelado a -20 °C, hasta su uso para la extracción del DNA.
Ganglios trigéminos de equinos. Para el estudio fueron extraídos los ganglios trigéminos de 38 de caballos sacrificados en una planta de faenado que cumple con las medidas mínimas necesarias para el sacrificio de equinos (según resolución 0222 del Ministerio de Salud). Luego del sacrificio de los animales, las cabezas fueron recolectadas y abiertas mecánicamente para extraer los ganglios trigéminos, los cuales fueron transportados al laboratorio en PBS de manera refrigerada. Una vez en allí, los ganglios fueron lavaron tres veces con PBS y se congelaron a -20 °C, hasta el momento de extraer el DNA.
Extracción del DNA de MNSP y ganglios trigéminos. Los MNSP fueron resuspendidos en 0.5 ml de tampón de lisis (10 mM Tris-HCl, pH 8.0; 100 mM EDTA; 0.5% SDS) e incubados en presencia de 5 μl de proteinasa K (20 μg/ml, Fermentas Inc® Cromwell Park Drive, USA) a 50 °C/3 h. El DNA se extrajo del lisado celular usando 0.5 ml de fenol: cloroformo: alcohol isoamílico (25:24:1), seguido por precipitación con 50 μl de acetato de sodio (3M) y 1 ml de isopropanol 100% (previamente enfriado a -20 °C). El DNA fue recolectado por centrifugación, se lavó dos veces con etanol al 70% y se disolvió en 200μl de tampón TE (Tris 10 mM, EDTA 1 mM).
Para la extracción del DNA de los ganglios trigéminos, se tomaron 150 mg de tejido y se cortaron en pequeños fragmentos; luego se sonicaron a 20 Kz/15 seg/hielo (este procedimiento se repitió tres veces). Finalmente, el DNA se extrajo usando el estuche comercial DNeasy Tissue (Qiagen Inc., Valencia, CA): en resumen, 100 μl del producto sonicado fueron lisados con 180 μl de tampón de lisis y 20 μl de proteinasa K; el lisado se depositó en una columna que contenía una membrana de gel de sílica (con afinidad por el DNA); la columna se lavó dos veces con 500 μl de tampón y finalmente el DNA fue eluido dos veces con 100 μl de tampón de elusión y almacenado a -20°C hasta su uso.
PCR semianidada para la gH de HVE-1 y la gB de HVE-4. Con el DNA obtenido de MNSP y de ganglio trigémino, se realizó una PCR semianidada para amplificar el gen que codifica por la glicoproteína H (gH) del HVE-1 y por la glicoproteína B (gB) de HVE-4, siguiendo el procedimiento reportado por Varraso et al (46); esta PCR es recomendada por la Organización Mundial para la Salud Animal (OIE) para diagnóstico de la infección por HVE en muestras de tejidos (2). La amplificación se realizó en un volumen final de 25 μl que contenía: tampón PCR 1X (50 mM KCl, 10 mM Tris/HCl, pH 9.0, 0.1% Tritón X-100), 200 μM de cada dinucleotido trifosfatado (dNTP), 2.5 mM MgCl2, 2.0 μM de cada cebador y 0.5 U de Taq DNA Polimerasa (Fermentas Inc® Cromwell Park Drive, USA). Para la gH de HVE-1 se utilizaron los cebadores Fw: 5’AAGAGGAGCACGTGTTGGAT3’ y Rw: 5’TTGAAGGACGAATAGGACGC3’; y para la gB de HVE-4 los cebadores Fw: 5’CTGCTGTCATTATGCAGGGA3’ y Rw: 5’CGTCTTCTCGAAGACGGGTA3’. Como control negativo se utilizó agua grado biología molecular (Amresco Inc® Solon, Ohio) en lugar de DNA. La amplificación se realizó en un termociclador (Perkin Elmer™ USA) con los siguientes parámetros: desnaturalización inicial a 95 °C /5 min, seguida de 35 ciclos de 95 °C/30 seg, 60 °C/30 seg y 72 °C/1 min, con una extensión final de 72 °C/5 min.
Para la segunda ronda de amplificación, se tomaron 2 μl del amplicón de la primera reacción (los cuales fueron usados como molde) y la nueva prueba se realizó bajo las mismas condiciones de la primera reacción, pero los cebadores Rw para HVE-1 y HVE-4 se reemplazaron por RN: 5’AGTAGGTCAGGCCGATGCTT 3’ y RN: 5’CGCTAGTGTCATCATCGTCG 3’, respectivamente. Finalmente, 5 μl del producto de la PCR semianidada fueron analizados por electroforesis en gel de agarosa al 2%, coloreado con bromuro de etidio (0.5 μM) y visualizados por trans-iluminación con luz UV.
Amplificación y secuenciación del ORF 68 y ORF 30 de HVE-1. Para amplificar el ORF 68 y ORF 30 de HVE-1, se realizó una PCR siguiendo las condiciones reportadas por Nugent et al (26). Los dos ORF se amplificaron de todas las muestras que fueron positivas para la gH del HVE-1. De cada muestra de DNA de MNSP y de ganglio trigémino se tomaron 3 μl y la reacción de amplificación se realizó en un volumen final de 50 μl, el cual contenía tampón PCR 1X (50 mM (NH4)2SO4, 10 mM Tris/HCl, pH 9.0, 0.1% Tritón X-100), 0.3 μM de cada uno de los cebadores descritos, 0.2 mM de cada dNTP, 1.5 mM MgCl2 y 1.25 U de Taq DNA polimerasa (Fermentas Inc® Cromwell Park Drive, USA). Para el ORF68 se utilizaron los cebadores ORF68 Fw: AGCATTGCCAAACAGTTCC y Rw: AAGAAACCACTGCTCAACC y para el ORF30 los cebadores fueron ORF 30 F: GCGCTACTTCTGAAAACG y R: CCACAAACTTGATAAACACG. Como control negativo se utilizó agua grado biología molecular (Amresco Inc® Solon, Ohio) en lugar de DNA. La PCR se llevó a cabo en un termociclador (Perkin Elmer™ USA) bajo las siguientes condiciones: una desnaturalización inicial a 94 °C/4 min, seguida por 35 ciclos de amplificación a 94 °C/30 seg, 48 °C/1 min y 72 °C/2 min, seguidos por una extensión final de 72 °C/10 minutos. Después de la amplificación, 5 μl de los productos de cada PCR fueron analizados en un gel de agarosa al 2%, coloreados con bromuro de etidio (0.5 μM) y visualizados por trans-iluminación con luz UV. Finalmente, la banda de interés fue cortada y purificada usando el estuche comercial “QIAquick PCR purification kit” (Qiagen Inc., Valencia, CA), siguiendo las instrucciones de la casa comercial. El DNA fue cuantificado usando un espectrofotómetro SmartSpect 3000™ (Bio-Rad, Hercules, California) a una longitud de onda de 260 nm y se envió a secuenciar.
Secuenciación y análisis de secuencias. Seis muestras de DNA aisladas de MNSP fueron enviadas a secuenciar a Macrogen® (Seul, Korea), siguiendo todas las recomendaciones establecidas por dicha casa comercial. La secuenciación se realizó en un secuenciador automático 3730xl DNA analyzer™, usando “ABI Prism BigDye™ Terminator Cycle Sequencing Ready Reaction Kit™” (Applied Biosystems® USA). La secuenciación se realizó en las dos direcciones con 15 μl (50 ng/μl) del amplicón purificado. Las secuencias fueron ensambladas y editadas usando el programa SeqMan™ (DNASTAR Inc., Madison, WI, USA). La secuencia de cada uno de los amplicones se comparó con secuencias de los genes correspondientes obtenidas del GenBank (números de acceso DQ180606 a 180738). Las secuencias de nucleótidos del ORF30 se utilizaron para realizar los análisis filogenéticos usando Mega® versión 3.1 (Center for Evolutionary Functional Genomics, Tempe, AZ, USA) (18) y BLAST pairwise alignments® en línea (NCBI, USA) (22) usando el método de mínima evolución.
]]> Análisis estadísticoLos resultados obtenidos se analizaron mediante estadística descriptiva, determinando promedios y desviación estándar y se realizó una prueba de Fisher, para evaluar si había diferencia estadística significativa (p<0.05) en la presentación de HVE-1 y HVE-4 en MNSP y en ganglios trigéminos. Todos los análisis se realizaron en los programas Prisma 4.0® (GraphPad Software, San Diego, CA, USA y Excel para Windows® (Microsoft®, USA).
Resultados
HVE -1 y HVE-4 infectan MNSP de equinos
Debido a que los HVE-1 y HVE-4 pueden producir una viremia asociada a células y puntualmente el HVE-1 puede inducir latencia en estas células, se tomaron los MNSP de equinos clínicamente sanos, de los cuales se conocía el estatus serológico para ambos virus. De los 96 caballos evaluados en Antioquia, se sabía que el 18.8% (18/96) presentaban anticuerpos contra la gG para HVE-1 y 98.7% (95/96) presentaban anticuerpos específicos para gG de HVE-4 (32). Por lo tanto, se tomó los MNSP de los individuos seropositivos y seronegativos (control negativo) para realizar una evaluación usando PCR semianidada.
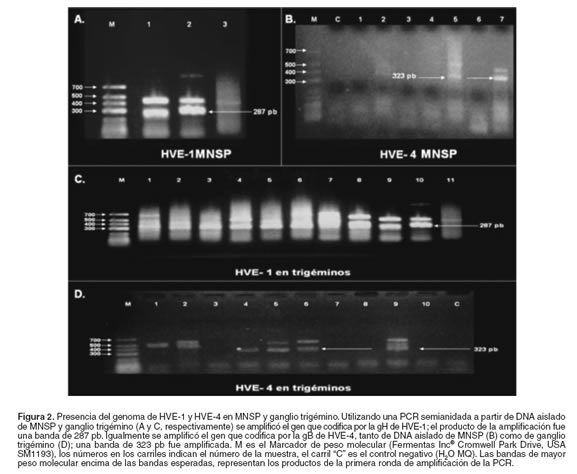
Según lo reportado por Varraso et al, mediante la PCR semianidada se obtienen bandas de 636 y 509 pares de bases (pb) para HVE-1 y HVE-4 respectivamente, después de la primera ronda de amplificación; y como resultado de la segunda ronda, se obtiene una banda de 287 y 323 pb, que corresponde al tamaño esperado para la secuencia blanco de los genes gH y gB de HVE-1 y HVE-4 (46). En efecto, como se aprecia en la figura 2, se lograron amplificar bandas del tamaño esperado en las muestras de DNA de MNSP y ganglios trigéminos. Sin embargo, como se puede observar en la misma imagen se evidencia la presencia de bandas de amplificación inespecífica de mayor tamaño al de la banda diagnóstica esperada; dichas bandas también son presentadas por los autores que reportaron la técnica (46); aunque no discuten su ocurrencia, en la práctica estas bandas inespecíficas pueden dificultar la toma de decisiones y conclusiones diagnósticas. Aún así, la técnica es aceptada y recomendada por la OIE para diagnóstico de HVE-1 y HVE-4 (2).
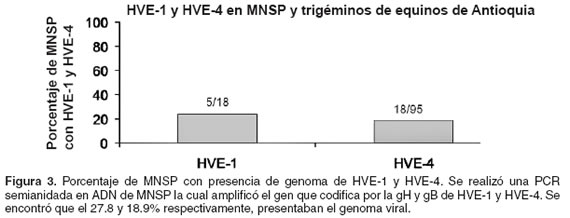
]]> Los resultados mostraron que de los 18 caballos que habían sido diagnosticados como seropositivos por la prueba de ELISA para el HVE-1, en cinco se logró amplificar el gen de la gH en MNSP, lo cual corresponde al 27.8% (véase Figura 3). Igualmente se encontró que de los 95 caballos que fueron seropositivos para HVE-4, por la prueba de ELISA, en 18 se logró amplificar el gen para la gB en dichas células, lo cual corresponde al 18.9% (véase Figura 3). Por el contrario, cuando se intentó amplificar el gen de las dos glicoproteínas a partir de DNA de MNSP de caballos que habían sido diagnosticados como seronegativos por la prueba de ELISA para ambos virus, no se obtuvo la banda esperada. Al comparar la frecuencia de detección del genoma de de HVE-1 y HVE-4 en los MNSP de los equinos, encontramos que no hay diferencia significativa (p>0.05) (véase Figura 3).
Infección por HVE-1 y HVE-4 en ganglio trigémino de equinos
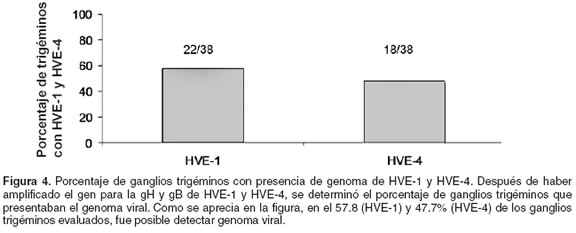
Para determinar la presencia de caballos reservorios de la infección por HVE-1 y HVE-4 en los caballos de Antioquia, se evaluó la presencia del genoma viral en ganglios trigéminos. Ante la imposibilidad para tomar biopsias de ganglio trigémino de los caballos en que se había encontrado anticuerpos para la gG de HVE-1 y HVE-4, se decidió realizar un muestreo post mortem en equinos no evaluados serológicamente de una planta local de faenado. Para el efecto se evaluaron un total de 38 ganglios trigéminos, en cuyas muestras se hallaron las bandas esperadas, pero también las bandas inespecíficas que se describieron en para los MNSP (véase Figura 2). Por su parte, la figura 4 muestra que en 22 del los 38 ganglios (58%) se amplificó el gen que codifica para la gH de HVE-1 y en 18 (48%) el gen para la gB de HVE-4.
Dado el alto grado de homología existente entre los genes de algunas de las principales glicoproteínas de HVE-1 y HVE-4 (gB, gC y gD) y considerando la existencia de reactividad antigénica cruzada para los dos virus (40) (25, 30, 47), se propuso corroborar la especificidad de la técnica; para esto los cebadores usados para amplificar la secuencias de la gB de HVE-4, se usaron para amplificar el gen de la gH de HVE-1. Tal como había sido reportado por los autores (46), no se logró ningún tipo de amplificación, lo que confirma la especificidad de la técnica.
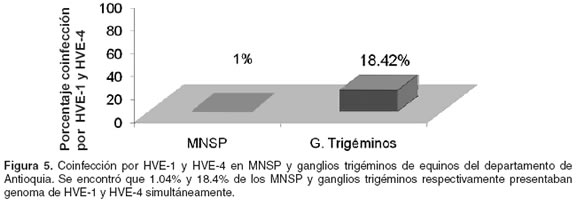
]]> Coinfección entre HVE-1 y HVE-4 en equinos del departamento de Antioquia Cuando se evaluó la coinfección en MNSP, uno de los 96 individuos muestreados (1.04 %) fue positivo en la amplificación tanto el gen de la gH de HVE-1, como del gB de HVE-4 (véase Figura 5). En ganglios trigéminos se amplificó el gen de las dos glicoproteínas en el 18.4% de los tejidos estudiados (véase Figura 5); es decir, de los 38 caballos evaluados, siete estaban coinfectados. Esto está indicando que posiblemente la coinfección es más frecuente en los ganglios trigéminos que en MNSP. Si bien se ha reportado que la coinfección es normal para estos dos HVE (14, 39), en el presente estudio es de gran importancia debido a que sirve como un indicativo de que los dos virus están presentes en la población de equinos del departamento de Antioquia.
Potencial neuropatogénico del HVE-1 circulante en el departamento de Antioquia.
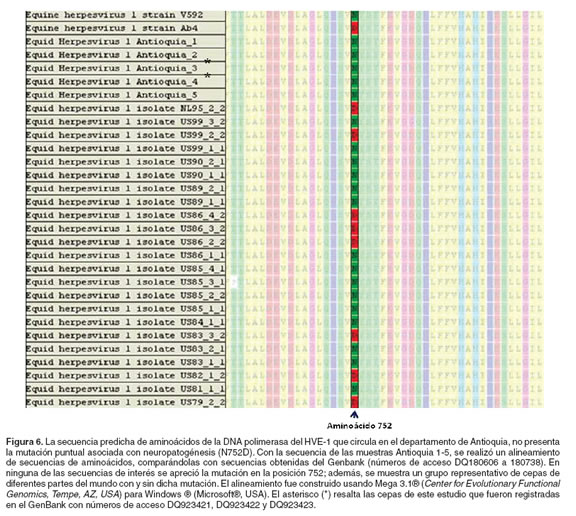
Debido a la falta de un aislamiento viral primario de la zona de estudio, que permitiera evaluar in vitro e in vivo la patogénesis de la infección, se decidió amplificar diferentes regiones del genoma viral, a partir de las muestras de DNA extraído de MNSP y de ganglios trigéminos que se encontraron positivas para la infección por HVE-1 por PCR semianidada. El ORF30 de HVE-1 se amplificó y secuenció, en el que se ha reportado una mutación puntual (cambio de aminoácido N752D, que corresponde con el cambio de nucleótidos G2254A), que tiene una fuerte asociación con neuropatogénesis; y el ORF68, el cual permite distinguir y categorizar los diferentes aislamientos virales, en seis grupos comunes, los cuales presentan gran restricción geográfica. La región del ORF30 amplificada corresponde a una región de 250 nucleótidos, localizada entre los nucleótidos 2200 a 2450 del genoma del HVE-1.
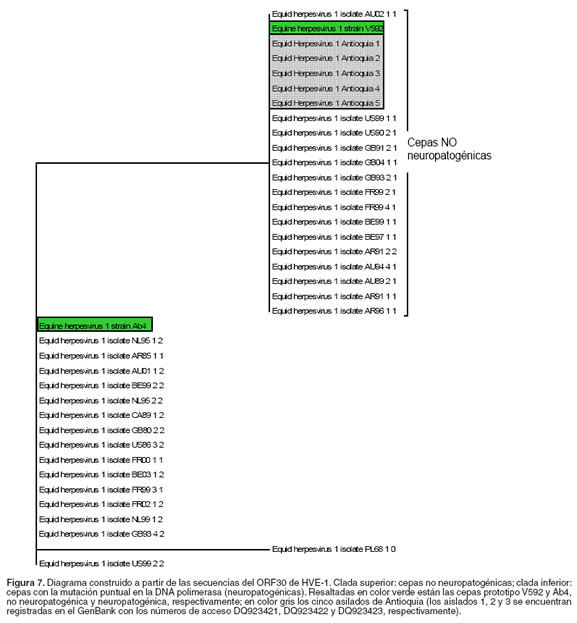
Las secuencias nucleotídicas del ORF30 obtenidas a partir de las muestras de DNA de MNSP fueron idénticas, tal como se aprecia en la figura 6, en las cinco secuencias de aminoácidos predichas obtenidas (Antioquia 1-5), no se encontró la mutación puntual (G2254A, correspondiente al cambio amiacídico N752D), en el gen de la DNA polimerasa; indicando que la cepa de HVE-1 que circula en el departamento de Antioquia, no está asociada con neuropatogénesis, al no poseer la mutación que se ha reportada asociada a este proceso patogénico del virus. En el presente trabajo no fue posible amplificar el ORF30 a partir de muestras de ganglio trigémino, debido quizás a la sensibilidad de la técnica. Al realizar la comparación de las secuencias Antioquia 1-5 con las diferentes secuencias del ORF30 reportadas en el GenBank, se encontró que la secuencia del ORF30 de las cepas pertenecientes al departamento de Antioquia es igual a la secuencia de dicho ORF de la cepa V592 (véase Figura 6, número de acceso AY464052), una cepa no neuropatogénica aislada a partir de un brote de enfermedad abortiva por HVE-1 (23, 24, 38); además, la secuencia de las cepas de Antioquia es igual a la secuencia de las cepas no asociadas con neuropatogénesis reportadas en el año 2006 (números de acceso DQ180606 a 180738), las cuales agrupan cepas aisladas de nueve países durante aproximadamente 30 años (26).
Luego, con los resultados obtenidos se realizó un análisis filogenético y se obtuvo una distribución en el gráfico de cladas, que aunque no es posible decir que se trata de un árbol filogenético propiamente dicho, permite observar cómo se agrupan las cepas con y sin la mencionada mutación y demuestra que los aislamientos de Antioquia (Antioquia 1-5) se ubican en la rama de las cepas no neuropatogénicas (véase Figura 7). Tres de las secuencias de los ORF30 obtenidas en este estudio (véase en Figuras 6 y 7, las cepas denominadas Antioquia 1, 2 y 3) fueron registradas en el Gen Bank con los números de acceso DQ923421, DQ923422 y DQ923423 respectivamente. Infortunadamente, las secuencias obtenidas con los amplificados del ORF68 de HVE-1, no fueron suficientes para poder determinar el posible origen del virus que circula en la región de Antioquia, debido a que las secuencias legibles en el electroferograma fueron demasiado cortas.
]]>
Discusión
La infección por HVE-1 y HVE-4 ha sido reportada en diferentes países del mundo. En Colombia, a pesar de poseer el reporte de un aislamiento viral (29), no se conocen estudios que confirmen la presencia de la infección por estos virus en la población de equinos. Estudios previos realizados por nuestro grupo de trabajo, mostraron evidencia serológica de la infección por HVE-1 y HVE-4 en dos departamentos de Colombia, por medio de una prueba de ELISA indirecta (32).
Si se tiene en cuenta que los caballos evaluados en la planta de faenado provenían de diferentes municipios del departamento de Antioquia (Colombia), los resultados presentados en este trabajo podrían sugerir que algún porcentaje de los caballos del departamento de Antioquia presentan infección latente con los HVE-1 y HVE-4. Similar a lo encontrado en MNSP, tampoco se encontraron diferencias estadísticamente significativas (p>0.05) en la detección de genoma viral de HVE-1 y HVE-4 en los ganglios trigéminos; es decir, posiblemente ambos virus tienen el mismo potencial para establecer infección latente en ganglios trigéminos de los equinos.
A partir de DNA aislado de MNSP de los caballos diagnosticados como seropositivos se logró amplificar el gen de la gH para HVE- 1 (27.8%) y el gen de la gB para HVE-4 (18.9%). Esto es muy importante si se tiene en cuenta que los MNSP además de estar asociados con la viremia de ambos virus, son las células implicadas en la dispersión y transmisión del virus a las células endoteliales de los vasos sanguíneos de otros sistemas orgánicos, tales como el sistema nervioso central y el útero, ocasionando las diferentes presentaciones clínicas (11).
Llama la atención que en los animales evaluados exista presencia de genoma viral en MNSP, aún estando clínicamente sanos al momento de la toma de la muestra y sin historia reciente de enfermedad respiratoria. Habitualmente, 4 a 6 días posinfección, el virus hace viremia asociada a células (MNSP), la cual puede durar hasta el día 27 después de una primoinfección (21). En dicho periodo los individuos ya han mostrado una recuperación clínica de la infección y las partículas virales están viajando a otros sistemas orgánicos en el hospedero (37) o estableciendo una infección latente. Nuestros resultados sugieren que las células (MNSP) están infectadas de manera activa por los HVE-1 y HVE-4, o bien que estos virus se encuentran en estado latente; sin embargo, la técnica utilizada no permite diferenciar entre estas dos alternativas. Es interesante anotar que en un estudio previo (32) se mostró que el 18.8% (18/96) de los equinos evaluados presentaban anticuerpos para la glicoproteína G (gG) del HVE-1; de ese 18.8%, en el 27.8% (5/18) se logró amplificar el gen de la gH, en MNSP. Para el caso del HVE-4, del 98.7% de seropositivos en el estudio anterior, el 18.9% presentaron el genoma viral en los MNSP. Estos resultados están mostrando que no en todos los caballos que presentan anticuerpos para proteína virales en suero, es posible amplificar el genoma viral en los MNSP.
Al igual que la gran mayoría de los miembros de la subfamilia Alfaherpesvirinae, los virus HVE-1 y HVE-4 establecen latencia en células nerviosas (neuronas sensoriales), después de una infección primaria. En este tipo de células, la transcripción es restringida, sin presencia de partículas virales; es decir, la detección de genoma viral es un indicativo muy probable de la infección latente (12, 35). Si bien, los ganglios trigéminos utilizados en esta investigación no se obtuvieron de los mismos caballos en los que se evaluó presencia de anticuerpos por serología y de genoma viral en MNSP por PCR (por la imposibilidad de sacrificar los animales para acceder a este ganglio), un alto porcentaje de las muestras analizadas presentaron genoma viral de HVE-1 y HVE-4 (57.9 y 47.7%, respectivamente). Lo anterior sugiere que hay un porcentaje de caballos del departamento de Antioquia clínicamente sanos pero con el genoma del HVE-1 y el HVE-4 latente en los ganglios trigéminos; ante una situación de estrés severo (por manejo, enfermedad o mala nutrición), es posible que estos animales puedan reactivar la infección viral y convertirse en fuente de contaminación para otros equinos que entren en contacto con ellos.
El HVE-1 (4, 8, 35) a diferencia de HVE-4 (6, 28, 48), establece infección latente tanto en MNSP, como en ganglio trigémino (33); sin embargo, no es posible aseverar que los animales de los que se amplificó el gen de la gH de HVE-1 o el gen de la gB de HVE-4 tuviesen una infección latente, debido a que no se evaluó presencia de transcriptos asociados a latencia (LATs); los cuales son un indicador de infección latente (17). Por otra parte, el porcentaje de ganglios trigéminos que presentaron genoma viral es similar a los resultados de otros investigadores, los cuales mostraron que es posible encontrar infección latente desde un 60 a 80% de la población de caballos muestreados (12).
]]> De manera independiente de que el virus esté o no en estado latente, tanto en MNSP como en ganglio trigémino, el alto porcentaje de detección del genoma viral en los dos tipos de tejidos, es un argumento que sugiere que ambos virus están plenamente establecidos en la población estudiada y, por ende, los mismos equinos constituyen reservorios de la infección, con capacidad de infectar otros caballos susceptibles. Es importante resaltar que, tanto los MNSP como los ganglios trigéminos, están coinfectados con los dos tipos de HVE; dicha coinfección ya había sido descrita por otros autores (13, 39), tanto para herpesvirus patógenos de animales como de humanos (9, 41). Sin embargo, el porcentaje de coinfección es mucho mayor en los ganglios trigéminos (principal sitio de establecimiento de la infección latente) que en los MNSP (34). Teniendo en cuenta que ambos virus pueden ser reactivados bajo un evento inmunosupresor, es posible que los animales puedan llegar sufrir reactivación de uno o de ambos virus, en forma simultánea.En Colombia no es común el reporte de casos de mieloencefalitis compatibles con la enfermedad ocasionada por el HVE-1 (comunicación personal con el Dr. Rafael Villalobos, del sistema de vigilancia epidemiológica de encefalitis virales del Instituto Colombiano Agropecuario ICA-CEISA). Los resultados de la amplificación y secuenciación del ORF30 de HVE-1, además de validar los resultados obtenidos, demuestran que la cepa circulante no posee la mutación descrita en la DNA polimerasa (26) (N752D, véase Figura 6), lo que permiten postular que las cepas de HVE-1 circulantes en Antioquia no poseen potencial neuropatogénico, en concordancia con el bajo reporte de casos de mieloencefalitis equina en la zona de estudio.
En conclusión, estos resultados en los que se ha usado diferentes estrategias para determinar la presencia de la infección por HVE-1 y HVE-4 en equinos del departamento de Antioquia (Colombia), confirman la presencia del agente infeccioso en la zona de estudio y sugieren que es posible encontrar genoma viral tanto en MNSP como en ganglios trigéminos; además, que la cepa de HVE-1 circulante no tiene potencial neuropatogénico. En su conjunto, estos resultados sugieren que existen individuos infectados por los HVE-1 y HVE-4, los cuales pueden actuar como reservorios de infección para otros equinos susceptibles.
Agradecimientos
Los autores agradecen al Grupo de Inmunovirología-Biogénesis por el apoyo brindado durante la ejecución de este trabajo; al doctor Claudio Berrío (MV) por propiciar el trámite del permiso para el acceso a las instalaciones de la planta de faenado, y a la Dra. Cecilia Galosi por su apoyo científico incondicional.
Referencias
1. Allen GP. Epidemic disease caused by Equine Herpesvirus-1: recommendation for prevention and control. Equine Vet Educ 2002; 14:136-142. [ Links ]
2. Allen GP. Equine rhinopneumonitis. In: OIE (Ed). Manual of diagnostic tests and vaccines for terrestrial animals. 5th ed. pp. 707-716. Office International des Epizooties, París, 2004. [Texto pdf] [ Links ]
3. Allen GP, Breathnach CC. Quantification by real-time PCR of the magnitude and duration of leukocyte-associated viraemia in horses infected with neuropathogenic versus non-neuropathogenic strains of Equid herpesvirus-1. Equine Vet J 2006; 38:252-257. [ [ Links ]Abstract]
4. Banbura M, Chmielewska A, Tucholska A, Malicki K. Occurrence of equine herpes virus type-1 (EHV-1)-specific DNA sequences in peripheral blood leukocytes of horses. Med Weter 2000; 56:521-523. [ Links ]
5. Baxi MK, Efstathiou S, Lawrence G, Whalley JM, Slater JD, et al. The detection of latency-associated transcripts of equine herpesvirus 1 in ganglionic neurons. J Gen Virol 1995; 76:3113-3118. [ Links ]
6. Borchers K, Wolfinger U, Lawrenz B, Schellenbach A, Ludwig H. Equine herpesvirus 4 DNA in trigeminal ganglia of naturally infected horses detected by direct in situ PCR. J Gen Virol 1997; 78:1109-1114. [ Links ]
7. Borchers K, Wolfinger U, Ludwig H. Latency-associated transcripts of equine herpesvirus type 4 in trigeminal ganglia of naturally infected horses. J Gen Virol 1999; 80:2165-2171. [ Links ]
8. Chesters PM, Allsop R, Purewal A, Edington N. Detection of latency-associated transcripts of equid herpesvirus 1 in equine leukocytes but not in trigeminal ganglia. J Virol 1997; 71:3437-3443. [ Links ]
9. Dunowska M, Meers J, Wilks CR. Isolation of equine herpesvirus type 5 in New Zealand. N Z Vet J 1999; 47:44-46. [ Links ]
10. Dutta SK, Myrup AC. Infectious center assay of intracellular virus and infective virus titer for equine mononuclear cells infected in vivo and in vitro with equine herpesviruses. Can J Comp Med 1983; 47:64-69. [ Links ]
11. Edington N, Bridges CG, Patel JR. Endothelial cell infection and thrombosis in paralysis caused by equid herpesvirus-1: equine stroke. Arch Virol 1986; 90:111-124. [ Links ]
12. Edington N, Welch HM, Griffiths L. The prevalence of latent Equid herpesviruses in the tissues of 40 abattoir horses. Equine Vet J 1994; 26:140-142. [ Links ]
13. Galosi CM, Vila Roza MV, Oliva GA, Pecoraro MR, Echeverria MG, et al. A polymerase chain reaction for detection of equine herpesvirus-1 in routine diagnostic submissions of tissues from aborted foetuses. J Vet Med B Infect Dis Vet Public Health 2001; 48:341-346. [ Links ]
14. Galosi CM, Barbeito CG, Vila Roza MV, Cid de la Paz V, Ayala MA, et al. Argentine strain of equine herpesvirus 1 isolated from an aborted foetus shows low virulence in mouse respiratory and abortion models. Vet Microbiol 2004; 103:1-12. [ Links ]
15. Gibson JS, Slater JD, Awan AR, Field HJ. Pathogenesis of equine herpesvirus-1 in specific pathogen-free foals: primary and secondary infections and reactivation. Arch Virol 1992; 123:351-366. [ Links ]
16. Goodman LB, Loregian A, Perkins GA, Nugent J, Buckles EL, et al. A point mutation in a herpesvirus polymerase determines neuropathogenicity. PLoS Pathog 2007; 3:e160. [ Links ]
17. Jones, C. Alphaherpesvirus latency: its role in disease and survival of the virus in nature. Adv Virus Res 1998; 51:81-133. [ Links ]
18. Kumar S, Tamura K, Nei M. MEGA3: Integrated software for molecular evolutionary genetics analysis and sequence alignment. Brief Bioinform 2004; 5:150-163. [ Links ]
19. Kydd JH, Smith KC, Hannant D, Livesay GJ, Mumford JA. Distribution of equid herpesvirus-1 (EHV-1) in respiratory tract of ponies: implications for vaccination strategies. Equine Vet J 1994; 26:466-469. [ Links ]
20. Kydd JH, Smith KC, Hannant D, Livesay GJ, Mumford JA. Distribution of equid herpesvirus-1 (EHV-1) in respiratory tract associated lymphoid tissue: implications for cellular immunity. Equine Vet J 1994; 26:470-473. [ Links ]
21. McCartan CG, Russell MM, Wood JLN, Mumford JA. Clinical, serological and virological characteristics of an outbreak of paresis and neonatal foal disease due to equine herpesvirus-1 on a stud farm. Vet Rec 1995; 136:7-12. [ Links ]
22. McGinnis S, Madden TL. “BLAST: at the core of a powerful and diverse set of sequence analysis tools. Nucleic Acids Res 2004; 32:W20-W25. [ Links ]
23. Mumford JA, Rossdale PD, Jessett DM, Gann S, Ousey J, et al. Serological and virological investigations of an equid herpesvirus 1 (EHV1) abortion storm on a stud farm in 1985. J Reprod Fertil Suppl 1986; 35:509-518. [ Links ]
24. Mumford JA, Hannant D, Jessett DM, O'Neill T, Smith KC, et al. Abortigenic and neurological disease caused by experimental infection with equid herpesvirus-1. Equine infectious diseases VII. Proc 7th Int Conf Equ Inf Dis. Tokyo, 1994 pp 261-275. [ Links ]
25. Nicolson L, Cullinane AA, Onions DE. The nucleotide sequence of an equine herpesvirus 4 gene homologue of the herpes simplex virus 1 glycoprotein H gene. J Gen Virol 1990; 71:1793-1800. [ Links ]
26. Nugent J, Birch-Machin I, Smith KC, Mumford JA, Swann Z, et al. Analysis of equid herpesvirus type 1 strain variation reveals a point mutation of the DNA polymerase strongly associated with neuropathogenic versus non-neuropathogenic disease outbreaks. J Virol 2006; 80:4047-4060. [ Links ]
27. Patel JR, Edington N, Mumford JA. Variation in cellular tropism between isolates of equine herpesvirus-1 in foals. Arch Virol 1982; 74:41-54. [ Links ]
28. Pusterla N, Leutenegger CM, Wilson WD, Watson JL, Ferraro GL, et al. Equine herpesvirus-4 kinetics in peripheral blood leukocytes and nasopharyngeal secretions in foals using quantitative real-time TaqMan PCR. J Vet Diagn Invest 2005; 17:578-581. [ Links ]
29. Ramírez GC, Chaparro JJ, Vera VJ, Villamil LC, Romero JR. Primer aislamiento de herpesvirus equino en Colombia. Rev Colomb Cienc Pecu 2001; 14:73 sup. [ Links ]
30. Riggio MP, Cullinane AA, Onions DE. Identification and nucleotide sequence of the glycoprotein gB gene of equine herpesvirus 4. J Virol 1989; 63:1123-1133. [ Links ]
31. Roizmann B, Desrosiers RC, Fleckenstein B, Lopez C, Minson AC, et al. The family Herpesviridae: an update. The Herpesvirus Study Group of the International Committee on Taxonomy of Viruses. Arch Virol 1992; 123:425-449. [ Links ]
32. Ruiz-Saenz J, Goez YP, Urcuqui-Inchima S, Gongora A, Lopez-Herrera A. Evidencia serológica de la infección por herpesvirus equino tipos 1 y 4 en dos regiones de Colombia. Rev Colomb Cienc Pecu 2008; 21:251-258. [ Links ]
33. Scott JC, Dutta SK, Myrup AC. In vivo harboring of equine herpesvirus-1 in leukocyte populations and subpopulations and their quantitation from experimentally infected ponies. Am J Vet Res 1983; 44:1344-1348. [ Links ]
34. Slater JD, Borchers K, Thackray AM, Field HJ. The trigeminal ganglion is a location for equine herpesvirus 1 latency and reactivation in the horse. J Gen Virol 1994; 75:2007-2016. [ Links ]
35. Smith DJ, Iqbal J, Purewal A, Hamblin AS, Edington N. In vitro reactivation of latent equid herpesvirus-1 from CD5+/CD8+ leukocytes indirectly by IL-2 or chorionic gonadotrophin. J Gen Virol 1998; 79:2997-3004. [ Links ]
36. Smith DJ, Hamblin AS, Edington N. Infection of endothelial cells with equine herpesvirus-1 (EHV-1) occurs where there is activation of putative adhesion molecules: a mechanism for transfer of virus. Equine Vet J 2001; 33:138-142. [ Links ]
37. Smith KC, Borchers K. A study of the pathogenesis of equid herpesvirus-1 (EHV-1) abortion by DNA in-situ hybridization. J Comp Path 2001; 125:304-310. [ Links ]
38. Smith KC, Whitwell KE, Mumford JA, Hannant D, Blunden AS, et al. Virulence of the V592 isolate of equid herpesvirus-1 in ponies. J Comp Path 2000; 122:288-297. [ Links ]
39. Taouji S, Collobert C, Gicquel B, Sailleau C, Brisseau N, et al. Detection and isolation of equine herpesviruses 1 and 4 from horses in Normandy: an autopsy study of tissue distribution in relation to vaccination status. J Vet Med B Infect Dis Vet Public Health 2002; 49:394-399. [ Links ]
40. Telford EA, Watson MS, Perry J, Cullinane AA, Davison AJ. The DNA sequence of equine herpesvirus-4. J Gen Virol 1998; 79:1197-1203. [ Links ]
41. Theil D, Paripovic I, Derfuss T, Herberger S, Strupp M, et al. Dually infected (HSV-1/VZV) single neurons in human trigeminal ganglia. Ann Neurol 2003; 54:678-682. [ Links ]
42. Thein P, Brown K. Infection mit equinen herpesviren und manifestation am zentralnervensystem beim pferd. Tierärztl Praxis 1988; 16:295-302. [ Links ]
43. Van der Meulen KM, Nauwynck HJ, Buddaert W, Pensaert MB. Replication of equine herpesvirus type 1 in freshly isolated equine peripheral blood mononuclear cells and changes in susceptibility following mitogen stimulation. J Gen Virol 2000; 81:21-25. [ Links ]
44. Van der Meulen KM, Nauwynck HJ, Pensaert MB. Mitogen stimulation favours replication of equine herpesvirus 1 in equine blood mononuclear cells by inducing cell proliferation and formation of close intercellular contacts. J Gen Virol 2001; 82:1951-1957. [ Links ]
45. Van der Meulen KM, Nauwynck HJ, Pensaert MB. Increased susceptibility of peripheral blood mononuclear cells to equine herpes virus type 1 infection upon mitogen stimulation: a role of the cell cycle and of cell-to-cell transmission of the virus. Vet Microbiol 2002; 86:157-163. [ Links ]
46. Varrasso A, Dynon K, Ficorilli N, Hartley CA, Studdert MJ, et al. Identification of equine herpesviruses 1 and 4 by polymerase chain reaction. Aust Vet J 2001; 79:563-569. [ Links ]
47. Whalley JM, Robertson GR, Davison AJ. Analysis of the genome of equine herpesvirus type 1: arrangement of cleavage sites for restriction endonucleases EcoRI, BglII and BamHI. J Gen Virol 1981; 57:307-323. [ Links ]
48. Welch HM, Bridges CG, Lyon AM, Griffiths L, Edington N. Latent equid herpesviruses 1 and 4: detection and distinction using the polymerase chain reaction and co-cultivation from lymphoid tissues. J Gen Virol 1992; 73:261-268. [ Links ]
¶ Para citar este artículo: Ruíz Sáenz J., Góez Y, López Herrera A. Detección de DNA de herpesvirus equino tipos 1 y 4 en mononucleares de sangre periférica y ganglio trigémino de equinos. Infección, latencia y una aproximación a la neuropatogénesis de la cepa circulante. Rev Colomb Cienc Pecu 2008; 21:372-386.
* Autor para el envío de la correspondencia y la solicitud de separatas. Dirección Actual: Grupo de Investigación en Microbiología y Epidemiología. Universidad Nacional de Colombia Sede Bogotá. Bogotá, Colombia. E-mail: julianruizsaenz@gmail.com
]]>