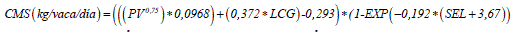
EFFECT OF INCREASING LEVELS OF DIETARY NONPROTEIN NITROGEN ON GLUCONEOGENIC PRECURSORS CONCENTRATION IN BOVINE LIVER
Rubén Darío Galvis Goez1; Diana Valencia Echavarría2; Héctor Jairo Correa Cardona3; Nicolás Fernando Ramírez Vásquez4 y Juliana Torres Sánchez5
1 Profesor Asociado. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. Departamento de Producción Animal. A.A. 1779, Medellín, Colombia. <rdgalvis@unalmed.edu.co>
2 Zootecnista. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <dmvalenc@unal.edu.co>
3 Profesor Asociado. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. Departamento de Producción Animal. A.A. 1779, Medellín, Colombia. <hjcorreac@unalmed.edu.co>
4 Profesor Asociado. Universidad de Antioquia. Facultad de Ciencias Agrarias. Escuela de Medicina Veterinaria, A.A. 1226, Medellín, Colombia. <nramirez@une.net.co>
5 Zootecnista. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <cjtorres@unal.edu.co>
Recibido: Septiembre 4 de 2009; Aceptado: Mayo 4 de 2010
Resumen. Con el fin de evaluar el efecto de niveles crecientes de proteína degradable en rumen (PDR) sobre las concentraciones de amonio y piruvato en hígado, se utilizaron 15 vacas Holstein que cursaban los primeros cien días posparto. Estas se asignaron aleatoriamente a tres tratamientos experimentales que consistieron en el suministro de un suplemento alimenticio comercial al que se le adicionó urea de tal manera que se incrementará el consumo de PDR en aproximadamente 0 (T0), 10 (T1) y 15% (T2) respecto a los requerimientos. A estas vacas se les determinaron las concentraciones hepáticas de amonio (Ahep) y piruvato (Pihep). Se realizó un balance nutricional de energía neta de lactancia (ENL), proteína metabolizable (PM), PDR y proteína no degradable en rumen (PNDR) donde se encontraron diferencias significativas entre los tratamientos (P<0,01) excepto para la ENL El balance para la PDR representó el 2,8, 9,5 y 13,1% de los requerimientos para T0, T1 y T2, respectivamente. Las concentraciones de Ahep difirieron entre tratamientos (P<0,06), mientras que no hubo diferencias respecto a las concentraciones de Pihep. Las variaciones en PM, PNDR y ENL fueron inversamente proporcionales con las de Ahep (P<0,04), mientras que las variaciones en Pihep fueron directamente proporcionales con las de Ahep (P<0,001). Todo lo anterior sugiere una alta utilización de la proteína metabolizable como fuente de energía, lo que condujo a exacerbar el exceso de nitrógeno y probablemente a restar aminoácidos para los procesos biosintéticos.
Palabras claves: Amonio, metabolismo del nitrógeno, precursores gluconeogénicos, proteína degradable en rumen.
Abstract. In order to test the effect of growing levels of diet rumen degradable protein on hepatic concentrations of ammonia and piruvate, fifteen cows Holstein in their first one hundred post calving days were used. They were assigned at random to three different experimental treatments, which consisted on the supply of a commercial feeding supplement without any addition of urea as a source of NNP (T0); the second treatment (T1) consisted on the supply of the same supplement with an addition of urea, so that the supply of PDR was increased 10% approximately to the requirements;finally, in the third treatment, (T2) the cows were given the same supplement which was added with enough urea so that the supply of PDR was increased 15% approximately to the requirements. In these cows were determined hepatic concentrations of ammonia (Ahep) and pyruvate (Pyhep). The concentrations of Ahep were different among treatments, whereas the concentrations of Pihep were similar. Negative relations between Ahep and ENL, Ahep and PNDR, Ahep, and PM, were found it; while Pihep was positively related with Ahep. These results suggest the metabolizable protein use as energy source, increasing nitrogen excess and probably rest amino acids for biosynthetic process.
Key words: Ammonia, gluconeogenic precursors, nitrogen metabolism, rumen degradable protein.
Los sistemas especializados de producción lechera en Colombia utilizan intensivamente la fertilización nitrogenada con la finalidad de generar aumentos significativos en la producción de forraje verde por hectárea. Esta fertilización; sin embargo, ocasiona cambios importantes en la calidad nutritiva de los forrajes afectando principalmente el contenido de las diferentes fracciones nitrogenadas y de los carbohidratos no estructurales (Carulla, 1999).
Las pasturas sobrefertilizadas pueden tener niveles bajos de carbohidratos fermentables (carbohidratos solubles (CS), fibra detergente neutra (FDN) y Pectina) al tiempo que incrementan la incorporación de N en la planta en forma de nitrógeno no proteico (NNP). Bajo estas condiciones el exceso de nitrógeno no puede ser utilizado por los microorganismos ruminales con lo que se satura la capacidad de utilización del amoniaco (NH3) por los microorganismos ruminales; así, el exceso de NH3 pasa a la sangre y es transportado hasta el hígado para ser convertido en urea (Correa et al., 2008). La formación de la urea trae consigo un trabajo metabólico que demanda sustratos y energía. En la síntesis de una mol de urea se gastan 3 moles de ATP y se utilizan (sin gasto aparente) metabolitos intermediarios como el oxaloacetato y el α-cetoglutarato. El papel principal de éstos radica en el transporte del amoniaco hasta el hígado, órgano en el cual se realiza la síntesis de urea (Kaneko, 1997).
El amonio que ingresa al ciclo de la urea es utilizado en primera instancia por la enzima carbamoil fosfato sintetasa I (E.C. 6.3.5.5; CPS- I), la afinidad de ésta enzima para el amonio es baja (Km=2 mmol/L) (Katz, 1992); por lo tanto la incorporación de amonio al ciclo de la urea por la CPS-I, es un paso limitante de la velocidad de formación de urea. Bajo condiciones de “represamiento” de amonio otras enzimas aumentan la captación de este sustrato; tal es el caso de la glutamato deshidrogenada que utiliza α-cetoglutarato y amonio para formar glutamato con una afinidad relativa mayor (Km=0,51-1,04 mmol/L) (Julliard et al., 1971). Ante la saturación del ciclo de la urea se detiene el ingreso de aspartato, la entrega de su respectivo grupo amino y la salida posterior de fumarato, lo que constituye una eliminación de un precursor gluconeogénico cuantitativamente importante. De esta forma la saturación del ciclo de la urea disminuiría la disponibilidad de precursores gluconeogénicos a través de la utilización de α-cetoglutarato en el transporte de amonio, explicando parcialmente, las bajas concentraciones de proteína en la leche observadas bajo las condiciones de producción que predominan en el trópico alto de Antioquia (Correa y Cuellar, 2004).
]]> Se conoce muy poco respecto al efecto de los niveles de amoníaco y/o urea sobre la actividad gluconeogénica bajo estas condiciones de producción. Aunque el ciclo de la urea esta sujeto a saturación, es de esperarse que los altos niveles de amoníaco presentes en el hepatocito alteren las condiciones químicas de éste, desfavoreciendo la actividad normal de la gluconeogénesis. En un estudio previo Galvis et al. (2003a) postularon que los excesos de proteína cruda dietaria (PCD) afectaban la tasa de gluconeogénesis disminuyendo la actividad de las enzimas claves de ésta vía; sin embargo, los resultados de este estudio no lo demostraron. Si los excesos de PCD no tienen efectos marcados sobre la actividad enzimática, es probable que los tenga sobre la tasa total de gluconeogénesis a través de la disminución en la disponibilidad de precursores. Es bien conocido que el ciclo de la urea, principal vía de detoxificación del amoniaco, utiliza gran cantidad de precursores gluconeogénicos, entre ellos α-cetoglutarato y también es bien conocido que la disponibilidad de éste limita la gluconeogénesis (Overton et al., 1998a).Según el marco teórico expuesto se postula que los excesos de nitrógeno no protéico dietario disminuyen la disponibilidad de precursores gluconeogénicos por la utilización de α-cetoacidos en la recolección de los excesos de amonio, por consiguiente el objetivo de esta investigación fue evidenciar el aumento en las concentraciones de amonio hepático y la disminución en las concentraciones de piruvato hepático conforme aumenta el consumo de nitrógeno no protéico.
MATERIALES Y MÉTODOS
Condiciones generales. El trabajo de campo se realizó durante los meses de diciembre de 2005 y enero de 2006, en el Centro Agropecuario Paysandú de la Universidad Nacional de Colombia, sede Medellín, ubicado a 2400 msnm, con una temperatura promedio de 14 ºC y una humedad relativa promedio de 80%.
Se utilizaron 15 vacas Holstein de segunda o más lactancias que cursaban los primeros cien días posparto. Las vacas fueron ordeñadas dos veces al día y diariamente se registró la producción de leche y la suplementación alimenticia de cada vaca. Adicionalmente, se utilizaron tres vacas secas canuladas en el rumen para la determinación de la degradabilidad de la proteína cruda de la dieta, utilizando el protocolo descrito y probado para forrajes tropicales por Giraldo et al. (1996). Los tiempos de incubación intraruminal fueron de: 0, 3, 6, 9, 12, 24, 48, 72, 84, y 96 horas, al final de los cuales se determino la degradabilidad de la proteína cruda (PC), siguiendo la metodología propuesta por Ørskov y Mc Donald (1979).
Forraje y dieta. Los animales pastoreaban praderas de kikuyo (Pennisetum clandestinum) sometidas al manejo tradicional con fertilización nitrogenada. Para la toma de muestras del forraje kikuyo, se realizo la técnica de “hand pluck” descrita por Euclides et al. (1962). Además, recibieron suplementación alimenticia acorde con la producción de leche y el número de lactancias. Las 15 vacas se asignaron aleatoriamente a tres tratamientos experimentales los cuales consistieron en el suministro de un suplemento alimenticio comercial sin adición de urea como fuente de NNP (T0); El segundo tratamiento (T1), consistió en el suministro del suplemento comercial al que se le adicionó urea de tal manera que se incrementará en 10% el consumo de proteína degradable en rumen (PDR) respecto a los requerimientos; finalmente, en el tercer tratamiento (T2) las vacas recibieron el suplemento comercial al que se le adicionó suficiente cantidad de urea para que el consumo de proteína degradable en rumen (PDR), fuera 15% mayor que los requerimientos.
El consumo de forraje se calculó como la diferencia entre la estimación del consumo total de materia seca y el consumo de materia seca del suplemento. Para la estimación del consumo total de materia seca (CMS) se utilizó la ecuación propuesta por el NRC (2001).
Toma de muestras. Al inicio del experimento las vacas fueron muestreadas con la finalidad de establecer las condiciones iniciales como covariables de la investigación. Luego fueron sometidas a las condiciones experimentales durante cuatro semanas consecutivas, al cabo de las cuales se realizó el muestreó final. Se tomaron biopsias hepáticas siguiendo el procedimiento descrito por Van den Top et al. (1995), con las que se obtuvieron muestras de aproximadamente 100 mg de tejido, las cuales se sumergieron en viales de 2 ml conteniendo una solución de suero fisiológico al 0,9% de NaCl. Todas las muestras se almacenaron a -10 ºC para su transporte y a -72 ºC hasta su análisis.
Caracterización nutricional de la dieta. Por otro lado, a las muestras de forraje y suplementos alimenticios se les determinó el contenido de cenizas
(Cen), extracto etéreo (EE), PC y proteína cruda insoluble en detergente ácido (PCIDA). La FDN)y la lignina se determinaron por el método de Van Soest (1985), utilizando acido sulfúrico al 72% para la determinación de lignina, mientras que el contenido de carbohidratos no estructurales se calculó por diferencia (NRC, 2001), utilizándolo como indicador para determinar la energía rápidamente fermentable y disponible en el medio ruminal.
Con los datos de los análisis químicos y degradabilidad, se calculó el contenido de energía neta de lactancia (ENL) del forraje y los suplementos utilizando las ecuaciones reportadas por el NRC (2001).
En un trabajo previo (Barrientos y Muñoz, 2006) a las muestras de forraje y suplemento alimenticio se les determinó la degradabilidad ruminal de la materia seca y de la proteína cruda.
La estimación del balance para ENL, PC, PDR y proteína no degradable en rumen (PNDR) se calculó por la diferencia entre los requerimientos estimados con el modelo NRC (2001) y los aportes de la dieta consumida por cada animal durante cada muestreo.
Determinación de amonio hepático. Para la determinación de amonio se utilizó el kit Aquamerck® 1.08024.0001 (Darmstadt R.F.A.), el cual es un kit cualitativo basado en la formación de un compuesto coloreado de azul de indofenol. Dado el carácter cualitativo del test original, se procedió a estandarizar la utilización del kit realizando la lectura calorimétrica en un espectrofotómetro Shimadzu UV-150-02. Para tal motivo se utilizó el procedimiento descrito por la casa comercial con algunas modificaciones, las cuales se describen a continuación: se tomaron 100 mL de sobrenandante del homogenado de hígado, 6 gotas del reactivo 1 (NaOH) y media micro cucharada (provista en el kit) del reactivo 2 (no identificado por la casa comercial) diluido en 2 mL de agua destilada, mezclándolos y dejándolos en reposo por 5 min; luego se adicionaron 2 gotas del reactivo 3 (nitroprusiato de sodio); se dejó en reposo por 7 min, para luego leer la absorbancia a 620 nm. Adicionalmente, se prepararon un blanco y una curva de calibración con concentraciones crecientes de cloruro de amonio (NH4Cl). Dada la dificultad para obtener el peso seco de las muestras de hígado, se procedió a cuantificar la proteína hepática presente en los sobrenadantes de los homogenados hepáticos por el método Bradford y utilizando el kit Protein Assay IIâ 500-0002 (Biorad, Richmond CA). Las concentraciones de amonio se expresaron como microgramos de amonio por microgramos de proteína hepática.
Determinación de piruvato hepático. En este trabajo el piruvato se estableció como un indicador de la disponibilidad de precursores gluconeogénicos. Para la determinación de piruvato se utilizó un ensayo enzimático acoplado a la caída en la absorbancia del NADH+H a 350 nm. La caída en la absorbancia a 350 nm es proporcional a las moles de NADH+H convertidas a NAD, y estas a su vez son proporcionales a las moles de piruvato convertido en lactato: a 1900 mL de un medio de reacción propuesto por Duff y Snell (1982) que contenía 100 mM-tris/HCl, 5 mM-MgCl2 20 mM de b - Mercaptoetanol y 0,15 mM de NADH+H, aun pH final de 7,5, se le adicionaron 50 mL de medio de reacción conteniendo 125 unidades de la enzima lactato deshidrogenasa (1.1.1.27) (Sigma L-2625) y 50 mL de sobrenadante de homogenado de hígado (muestra) e inmediatamente se registró la absorbancia a 350 nm. Cada tubo se incubó por 60 minutos en una incubadora Julabo TW 12, al cabo de los cuales la absorbancia dejó de disminuir. Posteriormente, se realizó una curva de calibración con concentraciones crecientes de NADH+H, para luego obtener las micromoles de piruvato presentes en las muestras. La concentración de piruvato se expresó como milimoles de piruvato por miligramo de proteína hepática.
Los análisis de metabolitos hepáticos se expresaron en relación a los miligramos de proteína hepática, dado que cuando se analizan biopsias de tejidos vivos, obtenidas de animales intactos de los cuales es imposible obtener grandes cantidades de muestra, que no puede ser sometida a tratamiento de secado para obtener el peso seco, es necesario cuantificar un marcador específico del tejido que varíe en la muestra de forma proporcional a la cantidad de tejido presente allí; para ello se utilizó la cuantificación de proteína hepática.
Análisis estadístico. Se realizó análisis de varianza para establecer las diferencias en cuanto a las concentraciones de amonio, y piruvato en tejido hepático, en función de los tratamientos. Cuando se presentaron diferencias estadísticamente significativas (P<0,05) se adelantó la prueba de Duncan para establecer las diferencias entre medias. Para ello se utilizó el PROC GLM del programa SAS (1998). Se utilizó el análisis de regresión para la exploración de las relaciones de asociación entre los metabolitos hepáticos amonio y piruvato con el balance de ENL, PDR, PNDR y proteína metabolizable (PM). La significancia estadística de la regresión se acepto cuando P<0,05; para lo cual se utilizó el PROC REG del programa estadístico SAS (1998).
]]>RESULTADOS
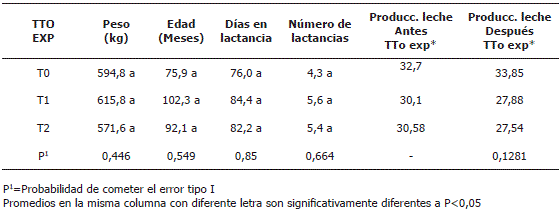
Características de los animales. La Tabla 1 presenta las principales características de los animales utilizados en la investigación en donde se puede apreciar que los grupos no difirieron significativamente en cuanto peso, edad y días en lactancia, lo que indica que respecto a estas variables la conformación de los grupos experimentales permitió la distribución aleatoria de los animales.
Tabla 1. Peso, días en lactancia, número de lactancias y edad de las vacas en experimentación.
Composición química y contenido energético de los alimentos. En la Tabla 2 se presenta la composición química del forraje y los suplementos experimentales. En esta se puede observar que la principal diferencia entre los suplementos alimenticios radicó en el contenido de PC, PDR y PNDR debido a la adición de urea. Es necesario aclarar que los tres suplementos fueron preparados teniendo como base un mismo lote de alimento concentrado, a partir del cual se prepararon los tratamientos experimentales por adición de urea; es decir, que cualquier diferencia en los valores de los parámetros composicionales diferentes a la PDR, son debidos a la variación normal dentro de una misma muestra. De igual manera, se observan diferencias entre los suplementos respecto al contenido de lignina que; sin embargo, no tuvieron efecto significativo sobre la valoración energética de los suplementos, ya que esta no presentó variaciones significativas entre tratamientos.
Tabla 2. Composición química y contenido energético de los alimentos utilizados en el estudio.
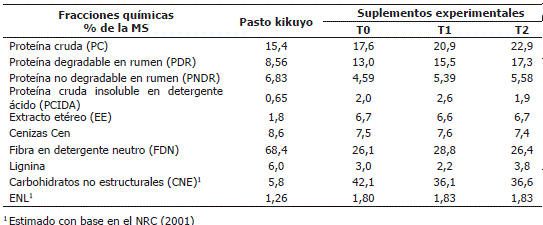
1 Estimado con base en el NRC (2001)
Balance nutricional. En la Tabla 3 se muestran los resultados de los balances nutricionales para los diferentes tratamientos. Como se puede apreciar se presentaron diferencias significativas (P<0,01) entre los tratamientos para los balances nutricionales, excepto para la ENL.
]]> Tabla 3. Promedio de los balances nutricionales para los diferentes tratamientos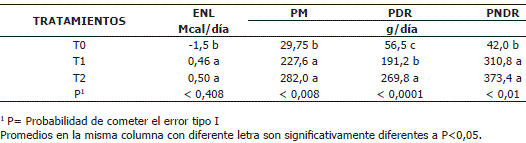
Es importante resaltar que el balance de PNDR principal factor condicionante en el diseño de este experimento, fue significativamente diferente entre tratamientos. La Tabla 4 informa acerca de las diferencias entre tratamientos para esta variable.
Tabla 4. Promedios de balance de proteína no degradable en rumen (PDR) expresados como porcentaje del requerimiento.

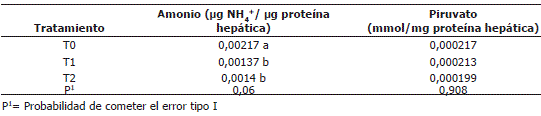
Amonio y piruvato hepáticos. Como se observa en la Tabla 5 hubo una tendencia a disminuirse el contenido de amonio hepático con la adición de urea en la dieta (P<0,06) mientras que los tratamientos no afectaron el contenido de piruvato hepático.
Tabla 5. Concentración de amonio y piruvato hepáticos.
Relaciones entre los balances nutricionales y las concentraciones hepáticas de piruvato y amonio. Como se observa en la Tabla 6, se encontró una relación directa entre las concentraciones de amonio y piruvato en hígado (P<0,01); De igual forma se encontraron relaciones inversas entre el balance de ENl y las concentraciones de amonio hepático y entre los balances de PNDR y de PM con las concentraciones hepáticas de amonio. Las demás relaciones propuestas en la Tabla 6 no presentaron significancia estadística.
Tabla 6. Relaciones entre los balances nutricionales y las concentraciones hepáticas de piruvato y amonio. ]]>

DISCUSIÓN
de los alimentos. El contenido de PC del pasto kikuyo fue relativamente bajo respecto a otros valores observados (Correa et al., 2008; Correa, 2006), mientras que el contenido de FDN (Correa, 2006) y lignina fueron más altos (Caro y Correa, 2006) lo que indica que los potreros se estaban pastoreando a edades de rebrote alta.
El contenido de ENL de este pasto se encontró dentro de los valores encontrados en el departamento de Antioquia (Correa et al., 2008); Sin embargo, es más alta que el valor observado por Caro y Correa (2006) para muestras de este pasto recolectadas a los 58 días de rebrote y con un menor contenido de FDN (62,88%). El contenido de lignina en las muestras analizadas por estos autores es más alto (7,93%) que el hallado en las muestras analizadas en el presente trabajo, siendo esta fracción la que marca la diferencia en cuanto a la digestibilidad de la FDN y, en consecuencia, al aporte que esta hace a la ENL.
Como era de esperarse, en la medida en que se incrementó la cantidad de urea adicionada al suplemento alimenticio, en esa medida se incrementó el contenido de PC, PDR y PNDR sin que se vieran afectadas las demás fracciones nutricionales. Respecto a la variación encontrada en los contenidos de lignina en los suplementos experimentales es necesario anotar que esta fue debida al método de muestreo, pues la composición básica de los suplementos fue la misma.
Balance nutricional. En concordancia con el planteamiento experimental, el balance para la PDR se incrementó en función de la adición de urea en los T1 y T2. Estos balances correspondieron al 2,75, 9,46 y 13,14% de los requerimientos para T0, T1 y T2, respectivamente (Tabla 4). Aunque T1 y T2 mostraron un mayor balance para la PM y la PNDR en comparación con el T0, estos no fueron diferentes entre sí debido a la variación en los datos (Tabla 4).
Amonio hepático. El grupo de vacas que presentó el valor más alto de amonio hepático (T0) también registró los balances más bajos de PM, PDR y PNDR, esto puede indicar que la elevación en los valores de amonio hepático no obedecieron al suministro dietario de fuentes proteicas, de tal modo que la elevación del amonio hepático en el tratamiento T0, posiblemente se debió a la utilización de aminoácidos corporales como fuente de energía, tanto para oxidación como para gluconeogénesis. Es así como Overton et al. (1998a, 1998b) sugieren que en vacas peri parturientas las cuales se caracterizan por un balance energético negativo (BEN), la tasa de gluconeogénesis a partir de alanina es más del doble comparada con la tasa observada en el preparto. Lo anterior tiene su sustentación dado que este grupo (grupo 1) fue el único que presento BEN, los restantes tratamientos presentaron un balance energético significativamente mayor y positivo. Por su parte Reynolds et al. (1994), afirman que la utilización de aminoácidos para gluconeogénesis da cuenta del 50% del amonio que se emplea para la síntesis de urea. De Visser et al. (1997), encontraron que el flujo de amonio desde el rumen a las vísceras que drenan al sistema porta (VDP) se incrementó con el nivel de fertilización nitrogenada, lo que era de esperarse debido al mayor consumo de PDR. En la presente investigación no se demostró un aumento en la concentración de amonio hepático acorde con el balance de PDR; estos resultados no coinciden por los observados por De Visser et al. (1997), dado que el flujo de amonio desde el rumen a VDP, no se corresponde exactamente con la concentración de amonio hepático puesto que la gran mayoría de amonio que llega al hígado es utilizado para la síntesis de urea. De este modo las concentraciones hepáticas de amonio ya han sufrido el efecto homeostático de este órgano. Symonds et al. (1981) encontró que el hígado de vacas lecheras fue capaz de remover amonio adicionado a la sangre portal hasta un suministro de 182 mg/min, pero a valores superiores a éste la concentración en sangre periférica se incrementa conforme aumenta su concentración en la vena porta.
La elevación en el balance de PDR no condujo a aumentos en la concentración ni de amonio plasmático ni de amonio hepático, por el contrario el grupo que tuvo un menor balance de PDR presentó los valores más altos de amonio hepático y esto no se correspondió con a la cantidad de amonio plasmático, lo que sugiere que la elevación de amonio hepático provino directamente de la desaminación oxidativa de aminoácidos, lo que reafirma lo expuesto anteriormente.
Piruvato hepático. El hecho de que las concentraciones de piruvato hepático no se hayan alterado con la elevación de PDR en T1 y T2, pudo ser debido a la continua remoción de piruvato para el mantenimiento de la gluconeogénesis, dada la necesidad constante de glucosa para el mantenimiento de la glucemia en un intervalo estrecho entre 40 a 60 mg/dL. Así, bajo condiciones nutricionales deficitarias en carbohidratos no estructurales como las que se presentan en el Centro Agropecuario Paysandú (Galvis, 2004) y que fueron las mismas encontradas en esta investigación, existen mínimas posibilidades para que se eleven las concentraciones hepáticas de piruvato. De Visser et al. (1997), encontraron que la liberación de lactato y propionato en VDP fue similar en las vacas que recibieron pasto Rye Grass (Lolium sp) con alto y bajo nivel de fertilización nitrogenada. Estos autores confirman la intensa tasa de gluconeogénesis a partir de propionato y en menor proporción a partir de aminoácidos y afirman que bajo condiciones deficitarias en almidones de sobrepaso puede ocurrir una utilización neta de glucosa por las VDP.
]]> Relaciones entre los balances nutricionales y las concentraciones hepáticas de piruvato y amonio. La relación directa entre las concentraciones de amonio y piruvato hepático, pudo ser debida a la continua remoción de los aminoácidos corporales para mantener la tasa de gluconeogénesis; así, todos los aminoácidos que están siendo utilizados como fuente de energía son desaminados elevando las concentraciones de amonio en el tejido hepático al tiempo que se genera una continua síntesis de piruvato. Lo anterior supone un aporte importante de carbonos de los aminoácidos hacia la síntesis de glucosa, debido a la continua utilización de glucosa por las vacas lactantes y a su disponibilidad limitada.La relación inversa entre las concentraciones de amonio hepático y el balance de ENL pudo ser debida a la utilización de aminoácidos corporales como fuente de energía, tanto para oxidación como para gluconeogénesis, ya que los animales que presentaron un balance de energía mayor necesitaron emplear aminoácidos como fuente de energía en menor cantidad que los animales con un balance energético menor. Como se observa en la Tabla 6, no se demostró un aumento de amonio hepático acorde con el nivel de NNP en la dieta, lo que reafirma que la elevación de los niveles de amonio fue debida al uso de estos como fuentes de carbonos para la gluconeogénesis y la oxidación. Lo anterior fue sugerido por Galvis et al. (2003b) quienes postularon que bajo las condiciones locales de producción, los aminoácidos son una importante fuente de carbonos para la formación de glucosa.
La relación inversa entre las concentraciones de amonio hepático y los balances de PNDR y PM pudo atribuirse al aporte neto de carbonos que realizaron las dietas con mayor cantidad de PNDR, al entregar estas mayor cantidad de proteína metabolizable y por ende de aminoácidos, los que al contrario de los aminoácidos de la reserva corporal llevan consigo un esqueleto carbonado que puede estar disponible para capturar otras moléculas de amonio ó para aportar intermediarios tanto al ciclo de la urea como al ciclo de Krebbs, vías metabólicas que directa e indirectamente disminuyen las concentraciones de amonio. Esto concuerda con la relación inversa entre balance de energía y amonio, pues las dietas con una mayor valoración energética presentaron en consecuencia un mayor aporte de proteína metabolizable.
Es necesario anotar que bajo las condiciones nutricionales imperantes en los sistemas de producción lechera del trópico alto colombiano, no es posible generar balances de PDR lo suficientemente bajos, como para que en una investigación se generen condiciones lo suficientemente contrastantes que permitan identificar con gran certeza sus efectos sobre la disponibilidad de compuestos intermediarios del metabolismo de la energía, como el piruvato.
De esta investigación se puede concluir que efectivamente existen relaciones entre el balance de proteína (PDR, PNDR y PM) y las concentraciones de metabolitos nitrogenados en tejido hepático. De igual manera se pudo observar que el balance de energía se relacionó negativamente con la concentración de amonio hepático, y que las concentraciones hepáticas de amonio y piruvato se relacionaron positivamente; todo lo anterior sugiere una alta utilización de la PM como fuente de energía, lo que conduce a exacerbar el exceso de nitrógeno y probablemente a restar aminoácidos para todos los procesos biosinteticos.
AGRADECIMIENTOS
A la Dirección de Investigaciones de la Universidad Nacional de Colombia, Sede Medellín por la financiación de esta investigación (código DIME: 30803874). Al personal operativo del Centro Agropecuario Paysandú por su apoyo y dedicación.
BIBLIOGRAFÍA
]]>Barrientos, S. y Y. Muñoz. 2006. Efecto de niveles crecientes de nitrogeno no proteico dietario sobre algunas variables metabólicas y nutricionales en vacas lactantes. Trabajo de grado. Facultad de Ciencias Agropecuarias. Universidad Nacional de Colombia. Medellín. 22 p. [ Links ]
Caro, F. y H.J. Correa. 2006. Digestibilidad posruminal aparente de la materia seca, la proteína cruda y cuatro macrominerales en el pasto kikuyo (Pennisetum clandestinum) cosechado a dos edades de rebrote. En: Livestock Research for Rural Development (18):10,http://www.cipav.org.co/lrrd/lrrd18/10/caro18143.htm; Consulta: noviembre 2008. [ Links ]
Carulla, J. 1999. Efectos de la fertilización nitrogenada sobre la proteína del forraje. En: Memorias. Simposio Internacional sobre la Proteína en la Leche. COLANTA. Medellín. 4 p. [ Links ]
Correa, H.J., M.L. Pabón y J.E. Carulla. 2008. Valor nutricional del pasto kikuyo (Pennisetum clandestinum Hoechst Ex Chiov.) para la producción de leche en Colombia (Una revisión): I. Composición química y digestibilidad ruminal y posruminal. En: Livestock Research for Rural Development (20): 59, http://www.cipav.org.co/lrrd/lrrd18/10/caro18143.htm; Consulta: noviembre 2008. [ Links ]
Correa, H.J. y A.E. Cuellar. 2004. Aspectos claves del ciclo de la urea con relación al metabolismo energético y proteico en vacas lactantes. Revista Colombiana de Ciencias Pecuarias 17(1): 29-38. [ Links ]
Correa, H.J. 2006. Posibles factores nutricionales, alimenticios y metabólicos que limitan el uso del nitrógeno en la síntesis de proteínas lácteas en hatos lecheros de Antioquia. En: Livestock Research for Rural Development 18: (3), http://www.cipav.org.co/lrrd/lrrd18/10/caro18143.htm; Consulta: noviembre 2008. [ Links ]
De Visser, H., H. Valk, A. Klop, J. Van Der Meuelen, J.G.M. Bakker and G.B. Huntington. 1997. Nutrient fluxes in splanchnic tissue of dairy cows: Influence of grass quality. Journal of Dairy Science 80(8): 1666–1673.
Duff, D.A. and K. Snell. 1982 Limitations of commonly used spectrophotometric assay methods for phosphoenolpyruvate carboxykinase activity in crude extract of muscle. Biochemistry Journal 206: 147-52. [ Links ]
Euclides, V., M. Macedo e M. Oliveira. 1962. Avaliacaõ de diferentes métodos de amostragen sob pastejo. Revista Sociedade Brasileira de Zooctecnia 21: 691-703. [ Links ]
Galvis, R.D. 2004. Consideraciones acerca de la sostenibilidad fisiológica de la vaca de alta producción lechera. Revista Colombiana de Ciencias Pecuarias 17(3): 290-297. [ Links ]
Galvis, R.D., H.J. Correa, N.F. Ramírez y W. Soler. 2003a. Influencia de las alteraciones metabólicas sobre la actividad PEPCK, la generación de IGF-1 plasmático y la reactivación ovárica en vacas en la lactancia temprana. Revista Colombiana de Ciencias Pecuarias 16(3): 228–236.
Galvis, R.D., H.J. Correa y N. Ramírez. 2003b. Interacciones entre el balance nutricional, los indicadores del metabolismo energético y proteico y las concentraciones plasmáticas de insulina, e IGF-1 en vacas en lactancia temprana. Revista Colombiana de Ciencias Pecuarias 16(3): 237-248. [ Links ]
Giraldo, L.A. 1996. Estandarización de la técnica de la biodegradación ruminal in situ, para evaluar forrajes tropicales. Revista Colombiana de Ciencias Pecuarias (Suplemento 9): 59-63. [ Links ]
Julliard, J.H., C. Godinot and C. Gautheron. 1971. Some modifications of the kinetic properties of bovine liver glutamate dehydrogenase (NADP) covalently bound to a solid matrix of collagen. Febs letters 14:185-188. [ Links ]
Kaneko, J.J., J.W. Harvey and M.L. Bruss. 1997. Clinical biochemistry of domestic animals. Fifth edition. Academic Press Inc, San Diego. 780 p. [ Links ]
Katz, N.R. 1992. Metabolic heterogeneity of hepatocytes across the liver acinus. The Journal of Nutrition 122(3 Suppl): 843-849. [ Links ]
Moller, S. 1996. “Protein penalty” in cows consuming pasture. Cattle Practice 4(1): 71.
National Research Council-NRC. 2001. The nutrient requirements of dairy cattle. Seventh revised edition. National Academy Press, Washington. 381 p. [ Links ]
Ørskov, E.R. and I. Mc.Donald. 1979. The estimation of protein degradation in the rumen from incubation measurements weighted according to rate of passage. The Journal of Agricultural Science 92: 499-503 [ Links ]
Overton, T.R., J.K. Drackley, C.J. Otteman-abbamonte, A.D. Beaulieu and J.H. Clark. 1998a. Metabolic adaptation to experimentally increased glucose demand in ruminants. Journal of Animal Science 76(11): 2938-46. [ Links ]
Overton, T.R., J.K. Drakley, G.N. Douglas, L.S. Emert, J.H. Clark. 1998b. Hepatic gluconeogenesis and whole body protein metabolism of periparturuent dairy cows as affected by sources of energy and intake of the prepartum diet. Journal of Dairy Science 81(suppl 1): 295. [ Links ]
Reynolds, C.K., D.L. Harmon and M.J. Cecava. 1994.Absorption and delivery of nutrients for milk protein synthesis by portal-drained viscera. Journal of Dairy Science 77(9): 2787-2808. [ Links ]
SAS/STAT. 1998. Users Guide: Statistics, Version 8. Cary, NC. [ Links ]
Symonds, H.W., D.L. Mather and K.A. Collis. 1981. The maximum capacity of the liver of the adult dairy cow to metabolize ammonia. British Journal of Nutrition 46: 481–86.
Van den Top, A.M., T.H. Wensing, M.J. Geelen, G.H. Wentink, A.T.H. Vant Klooster and A.C. Beynen. 1995. Time trends of plasma lipids and hepatic triacilglicerol synthesizing enzymes during postpartum fatty liver development in dairy cows with unlimited access to feed during dry period. Journal of Dairy Science 78:2208-2220. [ Links ]
Van Soest, P.J. and J.B. Robertson. 1985. Analysis of forrage and fibrous foods. A laboratory manual for animal science. Cornell University 503 p. [ Links ] ]]>