CARACTERIZACIÓN GENÉTICA DE AISLAMIENTOS DE PHYTOPHTHORA INFESTANS EN LAS ZONAS PRODUCTORAS DE PAPA DE LOS DEPARTAMENTOS DE ANTIOQUIA, BOYACÁ, CUNDINAMARCA Y NORTE DE SANTANDER (COLOMBIA)
GENETIC CHARACTERIZATION OF PHYTOPHTHORA INFESTANS ISOLATES IN THE POTATO-GROWING REGIONS OF ANTIOQUIA, BOYACA, CUNDINAMARCA AND NORTE DE SANTANDER (COLOMBIA)
Bernardo Silva1, 3; Sonia Jaramillo2,4; Mauricio Marín1, 5
1 Laboratorio de Biología Celular y Molecular, Fac. de Ciencias, Universidad Nacional de Colombia (Sede Medellín). Calle 59 A # 63-20. Medellín (Antioquia), Colombia.
2 Depto de Ciencias Agronómicas, Fac. de Ciencias Agropecuarias, Universidad Nacional de Colombia (Sede Medellín). Calle 59 A # 63-20. Medellín (Antioquia), Colombia.
]]> 3 bsilva0407@gmail.com4 sjaramal@gmail.com
5 mamarinm@unal.edu.co
Resumen
El tizón tardío ó gota de la papa, causada por el Oomycete Phytophthora infestans, es la enfermedad más limitante de este cultivo en el mundo. En Colombia, la gota presenta niveles altos de incidencia y severidad en la mayor parte de las regiones productoras de papa, siendo frecuente encontrar cultivos en los cuales toma proporciones epidémicas, causando la pérdida total de la producción cuando no se aplican frecuentemente fungicidas. El conocimiento de las características genotípicas de los fitopatógenos es una condición fundamental para el diseño y aplicación de métodos de control de las enfermedades causadas por éstos. En la presente investigación se evaluaron los haplotipos mitocondriales, tipo de apareamiento y el nivel de variación de las regiones ITS y 28S del ADN ribosomal de poblaciones de 40 aislamientos de P. infestans obtenidos de cultivos de papa en los departamentos de Antioquia, Boyacá, Cundinamarca y Norte de Santander (Colombia). Los resultados indicaron la presencia de un sólo haplotipo mitocondrial (IIa) y el tipo de apareamiento A1 en todos los aislamientos de las cuatro poblaciones evaluadas, lo que sugiere la ocurrencia de un solo linaje genético (posiblemente EC-1) del patógeno en los cultivos de papa de las zonas estudiadas. De otra parte, las secuencias de las regiones ITS y 28S fueron idénticas para todos los aislamientos y presentaron altos niveles de identidad con aislamientos de otros hospedantes y regiones del mundo. Estos resultados permiten inferir que en los cuatro departamentos las poblaciones de P. infestans presentan un comportamiento clonal y que no existe evidencia de su reproducción sexual.
Palabras clave: ARN ribosomal, ITS, gota de la papa, haplotipo mitocondrial, Phytophthora infestans, Solanum tuberosum
Abstract
Potato late blight caused by the Oomycete Phytophthora infestans is the most important disease of this crop world-wide. Late blight in Colombia presents high levels of incidence and severity in the potato-growing areas, taking epidemic proportions and causing a complete lost of production when fungicides are not applied at a frequent rate. Knowledge of genotypic characteristics of plant pathogens is a main condition to design and apply control measures for crop diseases. In this research, mitochondrial haplotypes, mating type and variation level of ITS and 28S ribosomal DNA sequences were evaluated in P. infestans populations of 40 isolates obtained in potato crops from the Colombian provinces of Antioquia, Boyaca, Cundinamarca, and Norte de Santander. Results indicated that all P. infestans isolates in the four populations showed the same mitochondrial haplotype (IIa) and had mating type A1, which suggests they possibly belong to the clonal lineage EC-1. Regarding ITS and 28S rDNA analysis, all Colombian isolates presented the same sequences for both regions and had high similarity with isolates from other hosts and regions of world. These results suggest that P. infestans populations attacking potato in four provinces of Colombia are mainly clonal and there is not evidence of their sexual reproduction.
]]> Key words: ITS, mitochondrial haplotype, Phytophthora infestans, potato late blight, ribosomal RNA, Solanum tuberosum
INTRODUCCIÓN
La gota de la papa es la enfermedad más devastadora de estos cultivos no sólo en Colombia sino también en otras regiones del mundo (Duncan 1999). Su agente causal es el Oomycete Phytophthora infestans (Mont.) de Bary (Peronosporales), el cual posee una gran plasticidad genética y altos niveles de virulencia sobre diversas plantas solanáceas. En Colombia, además de afectar papa, se han reportado daños en cultivos de tomate (Solanum esculentum), tomate de árbol (Solanum betaceum) y lulo (Solanum quitoense) en las zonas altoandinas del país. Ya que P. infestans es considerado como una amenaza para la seguridad alimentaria mundial, es uno de los fitopatógenos más estudiados en su reproducción, fisiología y procesos infectivos. Sin embargo, el conocimiento sobre los aspectos moleculares que subyacen su patogenicidad y los mecanismos de resistencia de plantas hospedantes y no hospedantes, así como su estructura poblacional en regiones como los Andes suramericanos, es aún incipiente.
Phytophthora infestans es un organismo heterotálico con dos tipos de apareamiento, A1 y A2, que se cree proceden de Toluca, México. Hasta el año de 1980, el tipo de apareamiento A2 estaba confinado a México, y se consideraba que solo el tipo A1 era el causante del tizón tardío en las demás regiones del mundo (Goodwin et al. 1994). Sin embargo, a partir de 1981 se presentaron reportes de la distribución del tipo de apareamiento A2 en varios países de Europa, Oriente medio, Asia y Sur América (Fry y Goodwin 1995); lo que ha conducido a que la población A1, haya sido desplazada por una nueva variante A1/A2. Dicha población ha resultado de la reproducción sexual de ambos tipos y se caracteriza por ser altamente virulenta y variable, lo cual le ha permitido adaptarse a nuevos hospedantes y a diferentes condiciones medioambientales (Fry et al. 1993, Fry y Goodwin 1995).
Alternativamente, existe una teoría que indica que el patógeno es originario de los Andes suramericanos, por ser la región de origen de sus principales hospedantes cultivados y silvestres (Abad et al. 1995, Gómez et al. 2007, Oyarzún et al. 1998). Sin embargo, la ausencia de reproducción sexual del patógeno en los Andes, según se desprende de estudios de variabilidad y flujo de genes, contradicen dicha hipótesis, aunque los hallazgos de ambos tipos de apareamiento sobre un mismo hospedante (S. muricatum) en Ecuador y la presencia de genotipos exóticos del Oomycete sobre hospedantes silvestres como S. ochrantum, S. hispidum y Brugmansia sanguinea han dado un nuevo auge a esta hipótesis (Adler et al. 2002a, b).
Morfológicamente P. infestans se caracteriza por presentar esporangios ovoides, elipsoides y limoniformes, semipapilados con pared delgada, y deciduos (Erwin y Ribeiro 1996). En la reproducción sexual de P. infestans participan anteridios anfígenos y oogonios, estructuras que deben corresponder a tipos de apareamiento diferentes, debido a la condición heterotálica del patógeno (Erwin y Ribeiro 1996). La producción de oosporas es estimulada por la presencia de sustancias hormonales tipo esterol. Estas esporas son típicamente estructuras de resistencia gracias a su fuerte pared y germinan por medio de un tubo a partir del cual se producen esporangios o directamente micelio (Agrios 1996, Wen-Hsiung 1998).
Tradicionalmente las poblaciones de P. infestans han sido caracterizadas con base en diferentes marcadores fenotípicos y genotípicos, destacándose el patrón electroforético generado a partir de la sonda RG57 mediante el empleo de la técnica de RFLP, el análisis de las isoenzimas Gpi y Pep, el nivel de sensibilidad al fungicida metalaxyl, la presencia de genes de virulencia determinados a partir de hospedantes diferenciales con genes R, la determinación del tipo de apareamiento y la evaluación de los haplotipos mitocondriales (Fry et al. 1991, Goodwin et al. 1994, Griffith y Shaw, 1998, Grunwald y Flier, 2005). El empleo de estas metodologías ha conducido a determinar que las poblaciones de P. infestans están subestructuradas a partir de linajes clonales, los que son frecuentemente denominados por las iniciales del país donde fueron encontrados y por un sufijo numérico que representa linajes derivados del original. Así por ejemplo, se considera que los aislamientos que participaron en la primera migración masiva de P. infestans desde México hacia Europa y EEUU, pertenecen al linaje US-1, mientras que aquellos reportados en Ecuador se denominan EC-1, EC-2 y EC-3 (Adler et al. 2004).
Actualmente los linajes de P. infestans que han sido reportados en Sudamérica que afectan con más frecuencia cultivos de papa, son aquellos con el patrón electroforético RG57 EC-1, cuyos aislamientos además se caracterizan por presentar tipo de apareamiento A1, perfiles isoenzimáticos Gpi 90/100 - Pep 96/100, y haplotipo mitocondrial IIa; mientras que en cultivos de tomate, el linaje corresponde al US-1 (tipo de apareamiento A1, Gpi 86/100 - Pep 92/100, y haplotipo mitocondrial Ib) y en tomate de árbol al EC-3 (tipo de apareamiento A1, Gpi 86/100 - Pep 76/100, y haplotipo mitocondrial Ia) (Adler et al. 2004, Mesa et al. 2008).
El estudio de la estructura metapoblacional de P. infestans en los Andes sudamericanos ha aportado interesantes resultados sobre el nivel de diversidad del patógeno. Así por ejemplo, en Ecuador se han descrito dos linajes EC-2 y EC-2.1 sobre solanáceas silvestres del complejo S. brevifolium (sección Amarrhichomenum) caracterizados por compartir los mismos patrones isoenzimáticos (Gpi 100/100 - Pep 76/100) pero que divergen en sus haplotipos mitocondriales y tipo de apareamiento: Ia - A1 y Ic - A2, respectivamente (Adler et al. 2004). Además se ha demostrado que la hipótesis de especificidad linaje clonal-hospedante no es absoluta, por cuanto sobre la especie cultivada S. muricatum (pepino de agua) se detectaron aislamientos de P. infestans pertenecientes tanto al linaje clonal US-1 como al EC-2.1 (Adler et al. 2004).
]]> Con respecto al grado de conocimiento del nivel de variación de P. infestans en Colombia, en un estudio que incluyó una población de 35 aislamientos de P. infestans obtenidos en diferentes hospedantes y regiones geográficas de Colombia, se determinó que todos los aislamientos pertenecen a los haplotipos mitocondriales Ia y IIa, los cuales están asociados con los linajes clonales EC-3 y EC-1, respectivamente. Además, la utilización de la técnica RAPD permitió determinar un grado de variación dentro de cada linaje, no explicable por la condición clonal del Oomycete en Colombia (Raigosa y Amaya 2006). Un resultado similar fue encontrado por Lagos (2002) quien al evaluar una población de 45 aislamientos de P. infestans obtenidos en diferentes municipios del departamento de Nariño, encontró que todos presentaban tipo de apareamiento A1 y haplotipo mitocondrial IIa. Más recientemente, Vargas et al. (2009) realizaron un estudio de caracterización de aislamientos de P. infestans de diferentes solanáceas del departamento de Cundinamarca y minoritariamente de Antioquia, encontrando un aislamiento con tipo de apareamiento A2 procedente de uchuva (Physalis peruviana), además de un muy bajo nivel de variación en la población analizada, la cual fue primordialmente asociada al linaje EC-1, aunque se encontraron tres variantes (COL-1, COL-2 y EC-1.1) con diferencias menores con respecto a este linaje. La importancia económica que representa P. infestans para la producción de papa en el país, hace necesario profundizar en el conocimiento de sus poblaciones con miras a generar mejores recomendaciones de manejo de la enfermedad de la gota de la papa. Esta investigación, se planteó con el objetivo principal de evaluar los niveles de variación de las poblaciones de P. infestans presentes en los cultivos de papa de los departamentos de Antioquia, Boyacá, Cundinamarca y Norte de Santander, mediante la definición de haplotipos mitocondriales, tipo de apareamiento y grado de divergencia de las secuencias de las regiones ITS y 28S del ADN ribosomal (ADNr).
MATERIALES Y MÉTODOS
Localización. Esta investigación se desarrolló en el Laboratorio de Biología Celular y Molecular de la Universidad Nacional de Colombia (Sede Medellín, Antioquia), Colombia. Para la obtención de una parte de los aislamientos bajo estudio, se realizó una colección de muestras sintomáticas de plantas de papa de diferentes variedades en los principales municipios cultivadores de los departamentos de Antioquia, Boyacá y Cundinamarca. Adicionalmente, se incluyeron aislamientos previamente colectados y almacenados en la Colección Nacional de Phytophthora infestans de la Universidad Nacional de Colombia (Sede Medellín) y cuyo origen corresponde a cultivos de estos tres departamentos y de Norte de Santander (anexo 1).
Aislamiento y purificación de P. infestans. Los aislamientos de P. infestans fueron obtenidos a partir de folíolos, porciones de tallo y pecíolos con síntomas de gota en cultivos de papa. Para el aislamiento del Oomycete se ubicaron los tejidos con síntomas sobre rodajas de papa variedad Tuquerreña (sin genes mayores de resistencia a P. infestans) con el fin de inducir el crecimiento de micelio, el cual se transfirió a medio agar-centeno (25 g de harina de centeno, 18 g de agar, 20 g de azúcar, 1 litro de agua destilada) y se conservó a 4 °C, incorporándose a la Colección Nacional de P. infestans.
Extracción de ADN. Los aislamientos de P. infestans fueron transferidos a medio líquido de arveja (25 g de arveja, 20 g de azúcar, 1 litro de agua destilada) e incubados a temperatura ambiente por 15 días, para filtrar el micelio, a partir del cual se extrajo el ADN siguiendo una modificación del protocolo del Centro Internacional de la papa (CIP 2000). Se procedió a la maceración de 100 mg de micelio con nitrógeno líquido, luego se adicionaron 500 μl de buffer de homogenización (0,1 M de TRIS HCL pH 8,0, 1,4 M de NaCl y 20 mM de EDTA pH 8,0) y se incubaron a 37 °C por 30 min. Después, se agregaron 500 μl de buffer de extracción CTAB 2X (CTAB 2% (p/v), 100 mM Tris HCl, pH 8,0, 1,4 M de NaCl, 20 mM EDTA pH 8,0) suplementado con 150 μl de SDS al 20% y se incubaron a 65 °C durante una hora. Posteriormente se agregaron 500 μl de fenol: cloroformo: alcohol isoamílico (25:24:1) y se mezclaron por inversión durante 15 min, para proceder a centrifugar a 13.000 rpm por 15 min. Al sobrenadante se le adicionaron 500 μl de cloroformo: alcohol isoamílico (24:1), se mezcló por inversión durante 10 min y se procedió a la centrifugación a 13.000 rpm por 10 min. El sobrenadante fue recuperado y se adicionaron 500 μl de alcohol isoamílico, agitando suavemente e incubándose una hora a -20 °C. El pellet generado fue lavado con etanol absoluto y luego resuspendido en 50 μl de agua destilada estéril y el ARN digerido mediante la adición de 5 μl de ARNasa [10 mg/ml] e icubado a 37 °C durante una hora. La integridad del ADN extraído fue determinada por medio de electroforesis en gel de agarosa al 0,8%, suplementado con 3 µl de bromuro de etidio [10 mg/ml] y utilizando como buffer de corrida TBE 0,5X (45 mM Tris-Borato, 1 mM EDTA pH 8). La visualización del ADN extraído se realizó en un transiluminador Bio Doc Analyze (Biometra, Göttingen, Alemania). La concentración de ADN fue calculada por absorbancia a 260 nm, utilizando un espectrofotómetro Thermo scientific, Genesys 6 (Waltham, EEUU), obteniéndose entre 115 y 232 ng/μl de ADN.
Determinación del tipo de apareamiento por PCR. Esta evaluación se realizó en 20 aislamientos de cada departamento, amplificando mediante PCR el arreglo S1 en el genoma de P. infestans (Judelson, 1996). Para este caso, el volumen final de cada reacción fue de 25 μl conformada por 0,32 µM de los iniciadores S1A (5'-AGGATTTCAACAA-3') y S1B (5'-TGCTTCCTAAGG-3'), 1X de buffer Judelson (20 mM Tris HCl pH 8,0; 10 mM KCl; 10 mM (NH4)2SO4; 30 mM MgSO4; 0.1% Triton X-100; dd H2O), 0,2 mM de deoxinucleótidos trifosfatos (dNTPs), 1 U de Taq ADN polimerasa (Fermentas, Vilnius, Lithuania) y 10-50 ng de ADN. Las amplificaciones se llevaron a cabo en un equipo PTC-100 (MJ Research, Whaltam, EEUU) con una desnaturalización inicial de 94 °C por 3 min; 40 ciclos de 94 °C por 30 s, 33 °C por 30 s, 72 °C por 60 s y una extensión final de 72 °C durante 3 min. Luego de la amplificación se tomaron 5 µl de los productos de reacción para analizarlos por electroforesis en gel de agarosa al 1,5% suplementado con 3 µl de bromuro de etidio [10 mg/ml]. La visualización de las bandas amplificadas se realizó bajo luz ultravioleta en un transiluminador automático Bio Doc Analyze (Biometra).
Determinación de haplotipos mitocondriales. Los haplotipos mitocondriales de los 160 aislamientos incluidos en el estudio fueron evaluados por la técnica PCR-RFLP usando los pares de iniciadores F1-R1 (F1: 5' GCAATGGGTAAATCGGCTCAA 3', R1: 5' AAACCATAAGGACCACA CAT 3'), F2-R2b (F2: 5' TTCCCTTTGTCCTCTACC GAT 3', R2b: 5'TTACGGCGGTTT AGCACATACA 3'), F3-R3 (F3: 5' ATGGTAGAGCGTGGGAATCAT 3', R3: 5' AAT ACCGCCTTTGGGTCCATT 3') y F4-R4 (F4: 5'-TGGTCATCCAGAGGTTTATGTT -3', R4: 5' CCGATACCGATACCAGCACCAA 3') (Griffith y Shaw 1998). Las condiciones del PCR fueron las siguientes: 25 μl de reacción total, 0,34 mM de cada par de iniciadores, 1X de buffer de PCR, 0,2 mM de dNTPs, 160 mg/ml de Albúmina de Suero Bovino (BSA), 1 U de Taq ADN polimerasa (Fermentas) y 10-50 ng de ADN. Las amplificaciones se llevaron a cabo en un equipo PTC-100 (MJ Research) y los ciclos de amplificación fueron los siguientes: desnaturalización inicial de 94 °C por 5 min; 40 ciclos de 94 °C por 30 s, 57 °C; 58 y 55 °C para los iniciadores F1-R1, F2-R2b, F3-R3 y F4-R4, respectivamente, por 60 s; 72 °C por 90 s, y una extensión final a 72 °C por 5 min.
Para la digestión, de cada amplificado se tomaron 5 μl y se mezclaron con 1 U de enzima, 1X de buffer de enzima y un volumen final de 20 μl con agua destilada estéril. Los amplicones obtenidos con el par de iniciadores F1-R1 fueron digeridos con la enzima CfoI, aquellos obtenidos con los iniciadores F2-R2b se digirieron con la enzima MspI y los amplificados con F3-R3 y F4-R4 se trataron con la enzima EcoRI. Las digestiones se realizaron al baño María a 37 °C durante 12 h y los resultados fueron visualizados en gel de agarosa al 2%, tal como se describió anteriormente.
Análisis de las regiones ITS y 28S del ADNr en aislamientos de P. infestans. Se tomaron al azar cinco aislamientos de cada una de las cuatro poblaciones estudiadas (anexo 1), a los cuales se les amplificó y secuenció la región ITS1, 5.8S e ITS2 mediante los iniciadores ITS4 (5' TCC TCC GCT TAT TGA TAT GC 3') e ITS5 (5' GGA AGT AAA AGT CGT AAC AAG G 3'), que permiten la obtención de un fragmento de 914 pb en Oomycetes (Cooke et al. 2000); mientras que para la región 28S se emplearon los iniciadores específicos para Oomycetes LROR (5' ACCCGCTGA ACTTAAGC 3') y LR6O (5' CGCCAGACGAGCTTACC 3') (Rietmüller et al. 2002) que amplifican una región del extremo 5' de la subunidad grande (28S) del ADNr, que contiene los dominios D1 y D2, ampliamente utilizados para estudios filogenéticos de hongos y Oomycetes (Guadet et al. 1989, Rietmüller et al. 2002). Las condiciones de PCR fueron similares a las descritas para la determinación de haplotipos mitocondriales, pero con temperaturas de alineamiento de 55 ó 52 °C para ITS y 28S, respectivamente.
]]> Una vez confirmado el tamaño de los amplicones, se procedió a su purificación mediante el kit QIAquick PCR Purification (Qiagen, CA, USA) para su secuenciación directa en ambas direcciones mediante el sistema Big Dye Terminator Cycle Sequencing Ready Reaction (PE Applied Biosystems, Foster City, EEUU) y su corrido en un secuenciador ABI Prism 3730XL (PE Applied Biosystems) de la compañía Macrogen (Corea del Sur).Las secuencias generadas con cada iniciador, se editaron mediante el software Chromas 1.45, deduciéndose las secuencias consenso y confirmándose su válidez por comparación con las bases de datos moleculares, mediante el programa BLAST (http://www.ncbi.nlm.nih.gov/BLAST/Blast.cgi). Posteriormente se procedió a su alineamiento con el algoritmo Clustal W incluido en el software Bioedit 6.0.6. De otra parte, se realizó un nuevo alineamiento utilizando secuencias correspondientes a aislamientos de P. infestans de diferentes regiones del mundo, así como también de las especies hermanas P. ipomoea, P. andina, P. mirabilis y P. phaseoli. Se utilizaron como grupos externos de análisis (outgroups) las secuencias de los Oomycetes P. nicotianae y P. palmivora, seleccionadas con base en el estudio filogenético realizado por Cooke et al. (2000).
El análisis filogenético se realizó utilizando el método de Máxima parsimonia (MP) con búsqueda heurística y TBR (tree-bisection-reconnection), mediante el software PAUP 4.0b10 (Swofford, 1998), realizando 1.000 remuestreos y tratando los espacios (gaps) como quinta base. El soporte de la topología interna de los dendrogramas fue evaluado mediante análisis de bootstrap con 1.000 remuestreos (Felsenstein 1985).
RESULTADOS
Aislamiento y purificación de P. infestans. En esta investigación se colectaron 151 muestras foliares de cultivos de papa con síntomas de gota. De éstas, 52 fueron obtenidas en cultivos de ocho municipios del departamento de Antioquia, mientras que 45 y 54 muestras procedían de 15 y 20 municipios de los departamentos de Boyacá y Cundinamarca, respectivamente. Una vez en el laboratorio, se logró la purificación de un total de 80 aislamientos, lo que corresponde a una eficiencia del 52,9%, la cual resulta bastante alta, ya que muchas muestras presentaban un alto nivel de residuos de fungicidas.
Se reactivaron otros 80 aislamientos de la Colección Nacional de P. infestans de la Universidad Nacional de Colombia sede Medellín, completando así un número de 160 cepas (anexo 1).
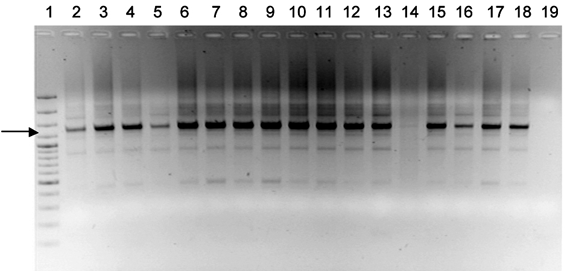
Determinación del tipo de apareamiento por PCR. En todos los casos, se logró la amplificación de un fragmento de 1.350 pb, que corresponde al fragmento del locus S1 de P. infestans, y que es indicativo del tipo de apareamiento A1 en este Oomycete (figura 1; anexo 1).
Figura 1. Resultado de la amplificación mediante PCR del locus S1, asociado al tipo de apareamiento A1 en Phytophthora infestans. El fragmento amplificado se indica con una flecha y corresponde a 1350 pb (Líneas: 1. Marcador de 100 pb. 2. BOY131. 3. BOY140. 4. BOY221. 5. BOY222. 6. BOY223 7. BOY224. 8. BOY225. 9. BOY226. 10. BOY227. 11. BOY228. 12. BOY229. 13. BOY230. 14. BOY235. 15. BOY101. 16. BOY236. 17. BOY237. 18. BOY238. 19. Control negativo)
]]>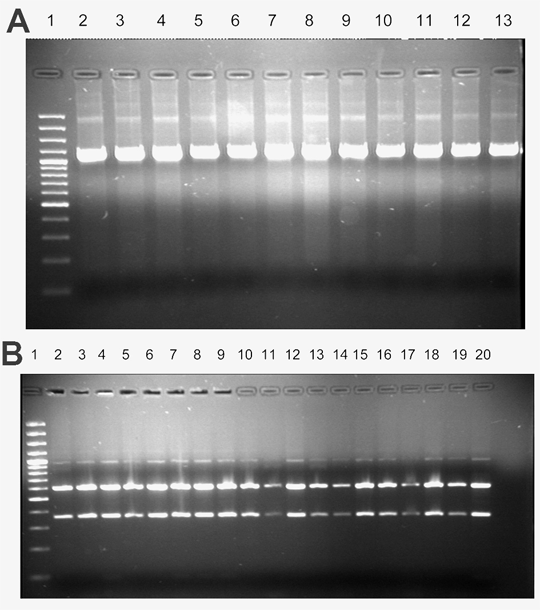
Figura 2. Digestión con las enzimas CfoI y EcoRI de los productos amplificados por PCR con los iniciadores F1-R1 (A) y F4-R4 (B) respectivamente, en diferentes aislamientos de Phytophthora infestans de Colombia, para la determinación de sus haplotipos mitocondriales (A = Línea 1. Marcador de 100 pb. 2. CUN123. 3. CUN139. 4. CUN148. 5. CUN112. 6. CUN105 7. CUN126. 8. CUN143. 9. CUN144. 10. CUN147. 11. CUN151. 12. CUN153. 13. CUN170. B = Línea 1. Marcador de 100 pb. 2. ANT13. 3. ANT18. 4. ANT19. 5. ANT25. 6. ANTU 7. ANT20. 8. ANT26. 9. ANT32. 10. ANT40. 11. ANT41. 12. ANT42. 13. ANT50. 14. ANT58. 15. ANT61. 16. ANT67. 17. ANT70. 18. ANT72. 19. ANT198. 20. ANT199)
Determinación de haplotipos mitocondriales. Los productos amplificados para cada par de iniciadores correspondieron a los previamente reportados por Griffith y Shaw (1998), con tamaños de bandas de 1.118 pb para F1-R1, 1.070 para F2-R2b, 1.308 pb para F3-R3 y 964 pb para F4-R4. Al realizarse la digestión de cada amplicón con las enzimas correspondientes, se encontró un único patrón de restricción para los 160 aislamientos evaluados, independientemente de la región y de la variedad de procedencia. De esta forma los fragmentos de restricción correspondieron a 1.118; 147-203-720; 1.308 y 361-603, para los productos F1-R1 (figura 2A), F2-R2b, F3-R3 y F4-R4 (figura 2B), respectivamente. Este resultado corresponde al haplotipo mitocondrial IIa (anexo 1), que de acuerdo con varias investigaciones está posiblemente asociado con el linaje clonal EC-1 (Adler et al. 2004, Jaramillo 2004).
Análisis de la región ITS del ADNr en aislamientos de P. infestans. Todas las secuencias de ITS de los 20 aislamientos de las zonas estudiadas, resultaron idénticas. Para el análisis filogenético, se incluyeron además de estas secuencias, nueve aislamientos de P. infestans de diferentes continentes y un representante de las especies hermanas: P. mirabilis, P. andina, P. ipomoea y P. phaseoli. Adicionalmente se utilizaron secuencias de P. palmivora y P. nicotianae como grupos externos (outgroups). El alineamiento final de las secuencias incluyó 811 sitios, de los cuales 680 caracteres fueron constantes, 110 variables pero no informativos y 21 resultaron informativos para el análisis de parsimonia. La longitud del árbol filogenético de MP fue de 140, con índices de consistencia IC = 0,99, retención IR = 0,91 y homoplasia IH = 0,014 (figura 3).
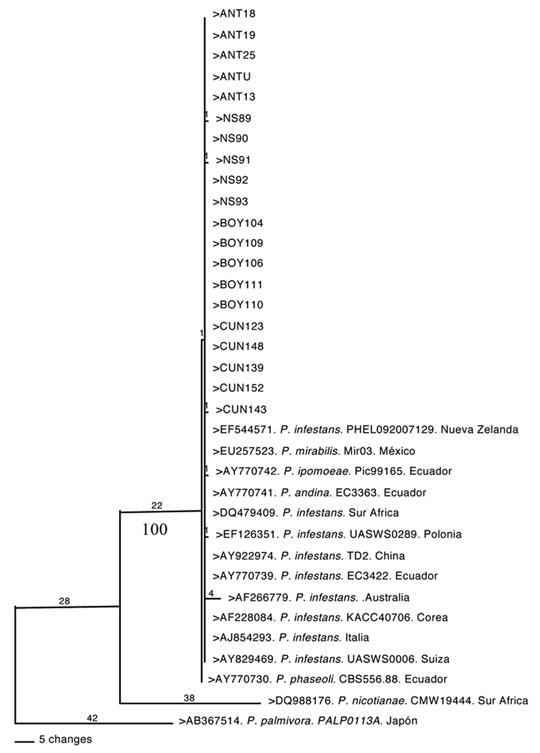
Figura 3. Árbol filogenético generado con secuencias de la región ITS del ADNr de aislamientos de Phytophthora infestans de cuatro departamentos de Colombia y otros países del mundo. El análisis de parsimonia incluyó secuencias de otras especies de Phytophthora con fines comparativos. Las diferencias en sitios de análisis se presentan sobre las ramas. Los valores de bootstrap = 50 se presentan bajo las ramas.
El análisis generó un árbol filogenético con un único clado y un soporte de bootstrap de 100%, que indistintamente incluye aislamientos de P. infestans, P. mirabilis, P. andina, P. ipomoea y P. phaseoli, los cuales compartieron secuencias idénticas con algunas excepciones puntuales, como aquella presente en el aislamiento de P. infestans de Australia.
Análisis de la región 28S del ADNr en aislamientos de P. infestans. Todas las secuencias de la región 28S en los 20 aislamientos de las cuatro poblaciones evaluadas resultaron idénticas. Para el análisis filogenético, se incluyeron además de estas secuencias, seis de los aislamientos de P. infestans de diferentes continentes y las secuencias de cada una de las especies hermanas P. andina, P. ipomoea y P. phaseoli. Adicionalmente, se utilizaron secuencias de P. palmivora y P. nicotianae como grupos externos. El alineamiento final de las secuencias incluyó 1.226 sitios, de los cuales 1.194 caracteres fueron constantes, 200 variables pero no informativos y 12 resultaron informativos para el análisis de parsimonia. La longitud del árbol filogenético de MP fue de 33, con IC = 0,97, IR = 0,92 e IH = 0,03 (figura 4). Al igual que el análisis de ITS, el árbol filogenético resultó con un sólo clado y un soporte de bootstrap del 100%, agrupando indistintamente aislamientos de P. infestans, P. andina, P. ipomoea y P. phaseoli, los cuales compartieron secuencias idénticas con la única excepción de un nucleótido en el representante de P. ipomoea.

DISCUSIÓN
Este trabajo representa un estudio genético detallado de P. infestans, el agente causal de la gota de la papa, que incluyó poblaciones en cultivos de papa de cuatro departamentos de Colombia (Antioquia, Cundinamarca, Boyacá y Norte de Santander), cuya área cultivada corresponde al 77% del total sembrado con este tubérculo en el país, con aproximadamente 122.617 ha (MADR 2006). Su desarrollo experimental se fundamentó en el empleo de diferentes técnicas moleculares incluyendo PCR específica para la definición de haplotipos mitocondriales y tipo de apareamiento, y secuenciación del ADNr (regiones ITS y 28S), metodologías que son ampliamente empleadas para este tipo de estudios (Cooke y Lees 2004, Gotoh et al. 2005) y por tanto constituyen un fuerte soporte para los resultados encontrados en la investigación.
Los resultados del análisis de haplotipos mitocondriales y tipo de apareamiento indican que las poblaciones del patógeno que afecta los cultivos de papa en los cuatro departamentos analizados, presentan un único haplotipo mitocondrial (IIa) y corresponden al tipo de apareamiento A1, lo que conduce a inferir su asociación con el linaje genético EC-1, frecuentemente encontrado en cultivos de papa de otros países andinos como Ecuador (Adler et al. 2004), Perú (Garry et al. 2005), Venezuela (Adler et al. 2007) y recientemente en Colombia sobre diferentes plantas solanáceas (Vargas et al. 2009). Sin embargo en dicho trabajo, Vargas et al. (2009) detectaron la presencia de tres aislamientos que presentaban diferencias menores en algunos de los caracteres evaluados, denominándolos como pertenecientes a los linajes COL-1 (diferencia en dos loci RFLP), COL-2 (diferencia en movilidad electroforética de Pep) y EC-1.1 (diferencia en un loci RFLP). Ya que dichos marcadores no se evaluaron en esta investigación, no es posible determinar la presencia de estas derivaciones del linaje EC-1 en la población bajo estudio. De interés resultó el hecho que no se detectó en ninguno de los 160 aislamientos analizados el haplotipo mitocondrial Ib (asociado al linaje US-1) que hasta hace algunos años se asociaba a algunos cultivos de papa del país (González y García 1998, Gilchrist 2001, Lagos 2002, León 2004). Esta situación apoya la hipótesis sobre el desplazamiento de poblaciones viejas por nuevas variantes introducidas o que se generan sexual ó parasexualmente y que presentan una mejor adaptación a las condiciones biológicas y medioambientales de los agroecosistemas. Así por ejemplo, Day y Shattock (1997) indican que la población de P. infestans presente en Inglaterra y Gales presentó un viraje total entre los años 1978 y 1995, al desaparecer el haplotipo Ib, siendo desplazado por aislamientos con haplotipos del tipo Ia y IIa; situación que se atribuye a la mayor agresividad de las nuevas variantes sobre los materiales vegetales de papa cultivados en estos países y a su resistencia al fungicida metalaxyl, lo cual representa una ventaja ecológica comparativa sobre las viejas poblaciones.
De otra parte, la ausencia de linajes genéticos reportados en otros países de Suramérica tales como el PE-3 (Perú; Garry et al. 2005), BR-1 (Bolivia, Brasil y Uruguay; Deahl et al. 2002, Adler et al. 2007) y AR-1 (Argentina; Adler et al. 2007), con los cuales Colombia tiene un intercambio comercial y migratorio muy activo, reafirman la necesidad de reforzar las medidas cuarentenarias del país para evitar el ingreso de patógenos en los tubérculos, a pesar de que el autoabastecimiento de papa en Colombia, conduce a que su importación sea mínima y se restrinja a fécula y papa procesada para la industria de alimentos (MADR 2006). Estas medidas cuarentenarias se hacen más necesarias, cuando se observa que todos los aislamientos de P. infestans evaluados en esta investigación (80) correspondieron exclusivamente al tipo de apareamiento A1, y que en muchos de los países de Suramérica es frecuente la ocurrencia de aislamientos del tipo A2 sobre papa, incluyendo Argentina, Brasil, Ecuador, Uruguay y Bolivia (Adler et al. 2007, Deahl et al. 2002, Reis et al. 2006); incluso se ha reportado la presencia de ambos tipos de apareamiento en los Andes ecuatorianos, aunque sobre diferentes hospedantes (Adler et al. 2002a, 2004) y de un aislamiento con el tipo de apareamiento A2 obtenido sobre plantas de uchuva en Colombia, aunque su patogenicidad en campo sobre variedades de papa no ha sido confirmada (Vargas et al. 2009). El éxito de las medidas fitosanitarias para evitar la introducción de genotipos A2 patogénicos en papa a nuevas regiones, tiene como ejemplo a Costa Rica, en donde desde 1970 se implementó un sistema nacional de certificación de semilla de papa y se establecieron exigentes regulaciones para la importación de semilla-tubérculo, manteniendo hasta ahora este país centroamericano libre del tipo de apareamiento A2 (Páez et al. 2004), a pesar de encontrarse geográficamente muy cerca de México, donde se registró por primera vez el tipo A2 y su ocurrencia es frecuente (Grunwald y Flier 2005).
La ausencia del tipo de apareamiento A2 en cultivos de papa en Colombia se ha reportado en los estudios basados tanto en las pruebas clásicas que utilizan cruces in vitro (González y García 1998, Gilchrist 2001, Lagos 2002) como en aquellas que emplean la técnica de PCR para la detección del locus S1 (Judelson 1996, León 2004). En esta investigación se empleó la técnica molecular de detección basada en PCR, resultado que fue confirmado por Marín et al. (2008) utilizando quince de los aislamientos (empleados en esta investigación) en evaluaciones in vitro con cruces entre todas las cepas y con la cepa de referencia A1, no siendo posible la observación de oosporas en ninguna de las combinaciones.
No se conoce la causa por la cual el tipo de apareamiento A2 en aislamientos de P. infestans de papa no se ha registrado en el país. Una posibilidad puede ser que se encuentre en otras solanáceas tal como ocurre en Ecuador sobre plantas del complejo S. brevifolium y S. muricatum (Adler et al. 2004) ó como se reportó recientemente en Colombia sobre plantas de uchuva (Vargas et al. 2009). También existe la posibilidad que el tipo A2 hubiese sido introducido al país desde otras latitudes, pero por las condiciones ambientales, de manejo y las características de las variedades de papa cultivadas, no se hubiera adaptado de manera exitosa. Adicionalmente, existe la posibilidad de que aislamientos del tipo A2 en papa esté en una proporción mínima dificultando su detección, pero que es suficiente para generar nuevas variantes del patógeno; ya se ha reportado que en países como Inglaterra la frecuencia del tipo A2 no supera el 3% (Day et al. 2004). En cualquier caso, la ausencia de uno de los tipos de apareamiento de un patógeno heterotálico hormonal como P. infestans en un cultivo, evita la reproducción sexual entre los miembros de sus poblaciones, situación que reduce la probabilidad de generación de variantes recombinantes, que tendrían ventajas agroecológicas como la capacidad de superar genes de resistencia en los materiales vegetales cultivados, la pérdida de sensibilidad a las moléculas fungicidas utilizadas para su control y la mejor adaptación a las condiciones medioambientales de las zonas productoras (Spielman et al. 1991, Fry y Goodwin 1995). Estas situaciones ya se han registrado en diferentes países en los cuales se han introducido ambos tipos de apareamiento. Así por ejemplo, se ha reportado que la estructura de las poblaciones europeas de P. infestans cambió significativamente después del ingreso de genotipos A2 a este continente a comienzos de los 80s, con aumento del grado de variabilidad genética de las poblaciones (Drenth et al. 1993), la diversidad de razas fisiológicas, el número de factores de virulencia del patógeno (Drenth et al. 1994) y el aumento de individuos con resistencia al fungicida metalaxyl (Day y Shattock 1997).
El análisis de secuencias de las regiones ITS y 28S del ADNr en el estudio, no mostró ninguna variación en dichas secuencias entre los veinte aislamientos evaluados (cinco representantes de cada departamento). De igual forma, el análisis filogenético indicó un muy bajo nivel de variación entre las secuencias evaluadas en los aislamientos colombianos y aquellas obtenidas de aislamientos de varios hospedantes de otras regiones del mundo, con excepción de un aislamiento de Australia, que en su secuencia ITS presentó cuatro sitios diferentes; aunque cabe la posibilidad que estas diferencias puedan deberse a errores en el análisis de las secuencias por parte de los depositantes en el GenBank. Dicho nivel de conservación en las secuencias ribosomales evaluadas, fue aún más evidente cuando se compararon con secuencias de ambas regiones en otras especies cercanas a P. infestans como P. andina, P. ipomoea, P. phaseoli y P. mirabilis (sólo región ITS), que igualmente fueron idénticas a las halladas en P. infestans.
Aunque no se esperaba un alto nivel de variación en el análisis de ITS entre los aislamientos Colombianos de P. infestans y aquellos de otras regiones, pues previamente Cooke et al. (2000) determinaron un alto nivel de similitud entre las secuencias de P. infestans y las de las especies P. phaseoli y P. mirabilis (clado 4 del árbol filogenético de Cooke et al. 2000), nuestra investigación buscaba determinar la presencia de algunas diferencias menores en las secuencias de esta región que pudieran ser útiles para el diseño de iniciadores específicos para el diagnóstico de los aislamientos colombianos del patógeno mediante PCR. Similarmente, esta búsqueda se realizó con base en los dominios D1-D2 de la región 28S del ADNr, que ha sido ampliamente utilizada para este tipo de aplicaciones en otros fitopatógenos del reino Chromista (Goker et al. 2003 en Peronosporales) y Fungi (Guadet 1989, en Fusarium sp.; Hople y Vilgalys 1999, en Coprinus sp.; Maier et al. 2003, en Uredinales), pero del mismo modo, las secuencias obtenidas en esta investigación fueron idénticas a las reportadas en el GenBank para aislamientos de P. infestans y de otras especies cercanas. Una alternativa para el hallazgo de sitios informativos con potencial para el diseño de iniciadores específicos corresponde a la secuenciación de genes funcionales. En este sentido Kroon et al. (2004) realizaron un análisis filogenético tendiente a diferenciar las especies del género Phytophthora utilizando dos genes mitocondriales (subunidad 1 del citocromo c oxidasa y subunidad 1 de la NADH deshidrogenada) y dos genes nucleares (factor de elongación 1a y ß-tubulina). Sus resultados permitieron la diferenciación de P. infestans de las especies cercanas antes mencionadas; sin embargo el uso de un aislamiento de cada haplotipo mitocondrial de P. infestans (Ia, Ib, IIa y IIb) no permite inferir si existía variación al interior de cada uno de ellos.
]]> Los métodos de diagnóstico molecular en P. infestans han sido previamente utilizados por varios investigadores y con diferentes propósitos. Así por ejemplo, Tooley et al. (1997) diseñaron los iniciadores PINF2, PERY2 y PNIC1 con base en secuencias ITS del ADNr para diferenciar las especies P. infestans, P. erythroseptica y P. nicotianae, que frecuentemente causan daños a los tubérculos de papa en los Estados Unidos. Sin embargo, de nuestro estudio es claro que esta región no permite la diferenciación de los aislamientos de P. infestans que afectan los cultivos de papa en Colombia. Más recientemente, Hussain et al. (2005), desarrollaron el par de iniciadores específicos INFFW2 e INFREV con base en esta misma región, con el fin de evaluar su utilidad en aplicaciones epidemiológicas como la cuantificación de inóculo en el suelo (oosporas y micelio) y para el diagnóstico asintomático de tubérculo-semilla, con muy buenos resultados. Existe una metodología no basada en análisis filogenético y genéricamente conocida como SCAR (sequence characterized amplified regions) para el diseño de iniciadores específicos de utilidad en el diagnóstico de patógenos y que pudiera explorarse en Colombia a partir de análisis con marcadores moleculares, como el desarrollado por Mesa et al. (2008) basado en AFLPs y que estudió los niveles de variabilidad de una población de P. infestans de cultivos de papa y tomate de árbol en el sur de Colombia. En este sentido recientemente Akino et al. (2008) diseñaron los iniciadores BD1 y BD2 a partir de la secuencia de un producto de RAPD denominado por los autores N605ab y que permite la separación de los linajes genéticos Japoneses de P. infestans, JP-1 del US-1 y de otros linajes japoneses (JP-2, JP-3 y JP-4). Se espera que esta aplicación sea igualmente incorporada por nuestro grupo, en trabajos futuros. Adicionalmente, es importante continuar con los estudios de diversidad genética del patógeno mediante marcadores moleculares, que permitan explorar una mayor extensión del genoma y así puedan soportar aún más la hipótesis sobre la condición clonal de las poblaciones de P. infestans que afectan la papa en Colombia.
AGRADECIMIENTOS
Esta investigación se realizó gracias al apoyo económico de la Vicerrectoría de investigaciones de la Universidad Nacional de Colombia a través del proyecto: 20101005571. La colección de los aislamientos utilizados en el estudio se ejecutó con el apoyo del personal técnico de Fedepapa y de los agricultores de Antioquia, Cundinamarca, Boyacá y Norte de Santander.
REFERENCIAS
1. Abad ZG, Abad JA, Ochoa C. 1995. Historical and scientific evidence that supports the modern theory of the Peruvian Andes as the centre of origin of Phytophthora infestans. En: Dowley LJ, Bannon E, Cooke RL, Keane T, Andosullivan E, editores. Phytopthora infestans 150. Dublin (Irlanda): Boole Press. p. 239-246. [ Links ]
2. Adler N, Chacon G, Forbes G, Flier W. 2002a. Phytophthora infestans sensu lato in South America population substructuring through host-specificity. Hamburgo (Alemania): Proceedings of the Global initiative on later blight conference. [ Links ]
3. Adler NE, Chacon G, Flier WG, Forbes GA. 2002b. The Andean fruit crop pear melon (Solanum muricatum), is a common host for A1 and A2 strains of Phytophthora infestans in Ecuador. Plant Pathology, 51: 802. [ Links ]
4. Adler N, Chacón G, Forbes G, Flier W [Internet]. 2007. Phytophtora infestans sensu lato in South America population substructuring through host-specificity. Fecha de consulta: Junio de 2008. Disponible en:http://library.wur.nl/file/wurpubs/LUWPUBRD_00347630_A502_001.pdf . [ Links ]
5. Adler NF, Erselius LJ, Chacón MG, Flier WG, Ordóñez ME, Kroon L, Forbes GA. 2004. Genetic diversity of Phytophthora infestans sensu lato in Ecuador provides new insight into the origin of this important plant pathogen. Phytopathology, 94: 154-162 [ Links ]
5. Agrios GN. 1996. Fitopatología. México D.F. (México): Limusa. [ Links ]
6. Akino S, Shirasawa Y, Kondo N, Naito S. 2008. N605ab, a specific molecular marker for Phytophthora infestans, distinguishes three genotypes in Japan. Journal of General Plant Pathology, 74: 125-127. [ Links ]
7. CIP (Centro Internacional de la Papa). 2000. Manual del curso ''Uso de marcadores moleculares en la caracterización de Phytophthora infestans (Mont.) de Bary''. Lima (Perú): Centro Internacional de la Papa. [ Links ]
8. Cooke DE, Lees AK. 2004. Markers, old and new, for examining Phytophthora infestans diversity. Plant Pathology, 53: 692-704. [ Links ]
9. Cooke D, Drenth A, Duncan J, Wagels G, Brasier CM. 2000. A molecular phylogeny of Phytophthora and related Oomycetes. Fungal Genetic and Biology, 30: 17-32. [ Links ]
10. Day JP, Shattock RC. 1997. Agressiveness and other factors relating to displacement of populations of Phytophthora infestans in England and Wales. European Journal of Plant Pathology, 103: 379-391. [ Links ]
11. Day, JP, Wattier R, Shaw D, Shattock R. 2004. Phenotypic and genotypic diversity in Phytophthora infestans on potato in Great Britain. Plant Pathology, 53: 303. [ Links ]
12. Deahl L, Cooke L, Black L, Wang T, Perez F, Moravec B, Quinn M, Jones R. 2002. Population changes in Phytophthora infestans in Taiwan associated with the appearance of resistance to metalaxyl. Pest Management Science, 58: 951-958. [ Links ]
13. Drenth A, Goodwin SB, Fry WE, Davidse LC. 1993. Genotypic diversity of Phytophthora infestans in the Netherlands revealed by DNA polymorphisms. Phytopathology, 83: 1087-1092. [ Links ]
14. Drenth A, Tas I, Govers F. 1994. DNA fingerprinting uncovers a new sexually reproducing population of Phytophthora infestans in The Netherlands. European Journal of Plant Pathology, 100: 97-108. [ Links ]
15. Duncan JM. 1999. Phytophthora an abiding threat to our crops. Microbiology Today, 26: 114-116. [ Links ]
16. Erwin DC, Ribeiro OK. 1996. Phytophthora diseases worldwide. St. Paul (Minnesota, USA): The American Phytopathological Society. [ Links ]
17. Felsenstein J. 1985. Confidence limits on phylogenies: an approach using the bootstrap. Evolution, 39: 783-791. [ Links ]
18. Fry EW, Drenth A, Spielman LJ, Mantel BC, Davidse LC, Goodwin S. 1991. Population genetic structure of Phytophthora infestans in the Netherlands. Phytopathology, 81: 1330-1336. [ Links ]
19. Fry EW, Goodwin SB, Dyer AT, Matuszak JM, Drenth A, Tooley PW, Sujkowski LS, Koh YJ, Cohen HA, Spielman JJ, Deahl KL, Inglis DA, Sandlan KP. 1993. Historical and recent migrations of Phytophthora infestans: Chronology, pathways, and implications. Plant Disease, 77: 653-661. [ Links ]
20. Fry EW, Goodwin SB. 1995. Recent migrations of Phytophthora infestans. En: Dowley LJ, Bannon E, Cooke RL, Keane T, Andosullivan E, editores. Phytopthora infestans 150. Dublin (Irlanda): Boole Press. Pp. 89-95. [ Links ]
21. Garry G, Forbes G, Salas A, Santa Cruz M, Pérez W, Nelson R. 2005. Genetic diversity and host differentiation among isolates of Phytophthora infestans from cultivated potato and wild solanaceous hosts in Peru. Plant Pathology, 54: 740-748. [ Links ]
22. Gilchrist E. 2001. Evaluación de la diversidad genética y patogénica de las poblaciones de Phytophthora infestans (Mont.) de Bary en Antioquia. [Tesis de Maestría en Biotecnología]. [Medellín, Colombia]: Facultad de Ciencias, Universidad Nacional de Colombia. [ Links ]
23. Goker M, Voglmayr A, Riethmuller M, Oberwinkler F. 2003. Taxomomic aspects of Peronosporaceae inferred from Bayesian molecular phylogenetics. Canadian Journal of Botany, 81: 672-683. [ Links ]
24. Gómez L, Carbone I, Ristaino JB. 2007. An Andean origin of Phytophthora infestans inferred from mitochondrial and nuclear gene genealogies. Proceedings of the National Academy of Science USA, 9: 3306-3311. [ Links ]
25. González C, García D. 1998. Caracterización de las poblaciones de Phytophthora infestans del altiplano cundiboyacense con base en el tipo de apareamiento y sensibilidad al fungicida metalaxyl. Fitopatología Colombiana, 22: 74-81. [ Links ]
26. Goodwin SB, Cohen BA, Fry WE. 1994. Pan global distribution of a single clonal lineage of the Irish potato famine fungus. Proceedings of the National Academy of Sciences USA, 91: 11591-11595. [ Links ]
27. Gotoh K, Akino S, Maeda A, Kondo N, Naito S, Kato M, Ogoshi A. 2005. Characterization of some Asian isolates of Phytophthora infestans. Plant Pathology, 54: 733-739. [ Links ]
28. Griffith GW, Shaw DS. 1998. Polymorphisms in Phytophthora infestans: Four mitochondrial haplotypes are detected after PCR amplification of DNA from pure cultures or from host lesions. Applied and Environmental Microbiology, 10: 4007-4014. [ Links ]
29. Grunwald N, Flier WG. 2005. The biology of Phytophthora infestans at its center of origin. Annual Review of Phytopathology, 43: 171-190. [ Links ]
30. Guadet J, Julien J, Francois J. 1989. Phylogeny of some Fusarium species, as determined by large-subunit rRNA sequence comparison. Molecular Biology Evolution, 6: 227-242. [ Links ]
31. Hopple JR, Vilgalys R. 1999. Phylogenetic relationships in the mushroom genus Coprinus and dark-spored allies based on sequence data from the nuclear gene coding for the large ribosomal subunit RNA: divergent domains, outgroups, and monophyly. Molecular Phylogenetics and Evolution, 13: 1-19. [ Links ]
32. Hussain S, Lees AK, Duncan JM, Cooke DE. 2005. Development of a species-specific and sensitive detection assay for Phytophthora infestans and its application for monitoring of inoculum in tubers and soil. Plant Pathology, 54: 373-382. [ Links ]
33. Jaramillo S. 2004. Monografía sobre Phytophthora infestans (Mont.) de Bary. Medellín (Colombia): Universidad Nacional de Colombia. [ Links ]
34. Judelson H. 1996. Genetic and physical variability at the mating type locus of the oomycete, Phytophthora infestans. Genetics, 144: 1005-1013. [ Links ]
35. Kroon LPN, Bakker FT., Van Den Bosch GB, Bonants P, Flier WG. 2004. Phylogenetic analysis of Phytophthora species based on mitochondrial and nuclear DNA sequences. Fungal Genetics and Biology, 41: 766-782. [ Links ]
36. Lagos LE. 2002. Aislamiento y caracterización genética de las poblaciones de Phytophthora infestans (Mont) de Bary en las zonas productoras de papa (Solanum tuberosum) en el departamento de Nariño. [Tesis de Maestría en Ciencias Biológicas]. [Cali, Colombia]: Facultad de Ciencias. Universidad del Valle. [ Links ] [ Links ]
38. MADR (Ministerio de Agricultura y Desarrollo Rural). 2006. Cadena productiva de la papa en Colombia. Fecha de consulta: Junio, 2008. Disponible en: http://www.minagricultura.gov.co. [ Links ]
39. Maier W, Begerow D, Wei M, Oberwinkler F. 2003. Phylogeny of the rust fungi: an approach using nuclear large subunit ribosomal DNA sequences. Canadian Journal of Botany, 81: 1-12. [ Links ]
40. Marín M. 2008. Informe final del proyecto de investigación: Variabilidad genética de aislamientos colombianos de Phytophthora infestans y su relación con el nivel de sensibilidad al fungicida del grupo de resistencia cruzada Qoi: fenamidone. Medellín (Colombia): Universidad Nacional de Colombia. [ Links ]
41. Mesa V, Mideros MF, Jaramillo S, Cotes JM, Lagos LE, Pineda R, Marin M. 2008. Variabilidad genética de aislamientos de Phytophthora infestans procedentes del suroeste de Colombia. Revista Iberoamericana de Micología, 25: 167-172. [ Links ]
42. Oyarzún PJ, Pozo A, Ordoñez ME, Doucett K, Forbes GA. 1998. Host specificity of Phytophthora infestans on tomato and potato in Ecuador. Phytopathology, 88: 265-271. [ Links ]
43. Páez O, Valverde R, Gómez L, Brenes A. 2004. Diversidad genética de aislamientos de Phytophthora infestans en plantaciones de papa en Costa Rica con el uso de RAPDs. Agronomía Costarricense, 29: 41-55. [ Links ]
44. Reis A, Fabiana HS, Ribeiro S, Eduardo SG, Mizubuti. 2006. Caracterização de isolados de Phytophthora infestans do Distrito Federal e de Goiás. Fitopatologia Brasileira, 31: 270-276. [ Links ]
45. Raigosa NA, Amaya MC. 2006. Análisis de la variabilidad genética de aislamientos Colombianos de Phytophthora infestans utilizando marcadores moleculares. [Trabajo de grado Ingeniería Agronómica]. [Medellín. Colombia]: Universidad Nacional de Colombia. [ Links ]
46. Riethmüller A, Voglmayr H, Göker M, Weiß M, Oberwinkler F. 2002. Phylogenetic relationships of the downy mildews (Peronosporales) and related groups based on nuclear large subunit ribosomal DNA sequences. Mycologia, 94: 834-849. [ Links ]
47. Spielman LJ, Drenth A, Davidse LC, Sujkowski LJ, Gu W, Tooley PW, Fry WE. 1991. A second world-wide migration and population displacement of Phytophthora infestans?. Plant Pathology, 40: 422-430. [ Links ]
48. Swofford DL. 1998. PAUP: Phylogenetic analysis using parsimony (* and other methods). Version: 4. Sunderland (USA): Sinauer Associates. [ Links ]
49. Tooley PW, Bunyard BA, Carras MM, Hatziloukas E. 1997. Development of PCR Primers from Internal Transcribed Spacer Region 2 for detection of Phytophthora species infecting potatoes. Applied and Environmental Microbiology, 63: 1467-1475. [ Links ]
50. Vargas, AM, Quesada-Ocampo LM, Céspedes MC, Carreño N, González A, Rojas A, Zuluaga AP, Myers K, Fry WE, Jiménez P, Bernal AJ, Restrepo S. 2009. Characterization of Phytophthora infestans populations in Colombia: First report of the A2 mating type. Phytopathology, 99: 82-88. [ Links ]
51.Wen-Hsiung KO. 1998. Chemical stimulation of sexual reproduction in Phytophthora infestans. Botanical Bulletin of Academia Sinica, 39: 81-86. [ Links ]
]]> ANEXOS
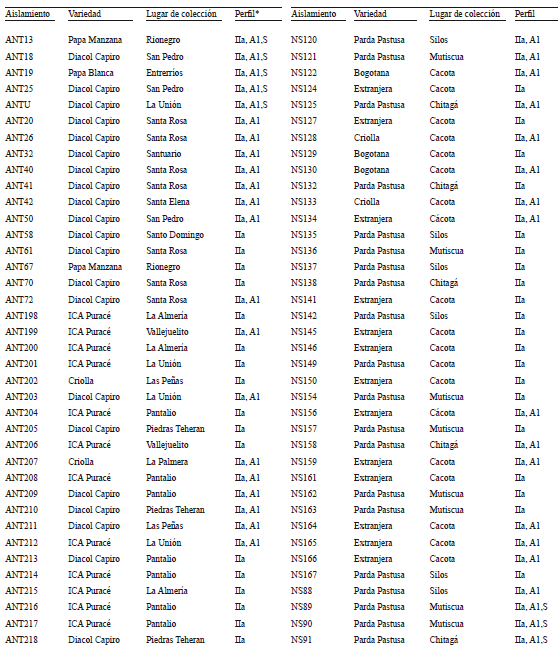
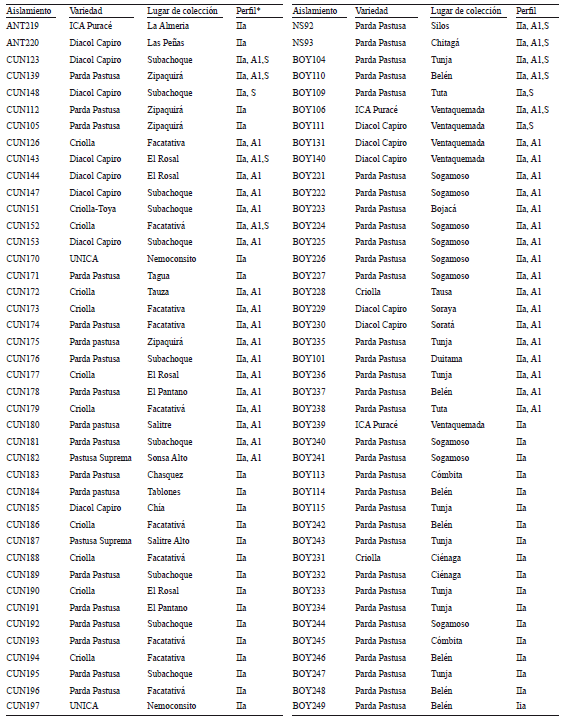
Anexo 1. Aislamientos de Phytophthora infestans procedentes de cultivos de papa (Solanum tuberosum y S. phureja) en Antioquia, Boyacá, Cundinamarca y Norte de Santander (Colombia) (* = Perfil se refiere al haplotipo mitocondrial, tipo de apareamiento y S a los aislamientos que fueron secuenciados para las regiones ITS y 28S del ADNr, cuyos números de accesión del GenBank corresponden a los rangos FJ869977- FJ869996 y FJ869957-FJ869976, respectivamente)
Recibido:enero 2009
]]> Aceptado: junio 2009 ]]>