PABLO LOZANO
RAINER BUSMANN
MANFRED KUPERS
DIEGO LOZANO C.
Universidad de Hohenheim, Instituto de Botánica, 2194 Garbenstr. 30, 70599 Stuttgart, Alemania. pablo_lozano@hotmail.com
]]> William L. Brown Center for Plant Genetic Resources, Missouri Botanical Garden, P.O. Box 299, St. Louis, MO 63166-0299, U.S.A. rainer.bussmann@mobot.orgUniversidad de Hohenheim, Instituto de Botánica, 2194 Garbenstr. 30, 70599 Stuttgart, Alemania.
Universidad Nacional de Loja, Herbario Reinaldo Espinosa (LOJA), Ecuador.
RESUMEN
El Parque Nacional Podocarpus (PNP), ubicado al sur del Ecuador, ocupa una posición climatológica clave con respecto a la deflexión de Huancabamba aledaña a esta área hacia el sur, que ocasiona una barrera natural de distribución de especies y por otro lado este aislamiento biogeográfíco ha originado especiación florística en algunas especies. La geografía accidentada de esta región, sumada a la escasa capa vegetal y precipitaciones que superan los 6000 mm en los páramos, ocasiona impactos severos en los flancos orientales y occidentales de la baja Cordillera Oriental que no sobrepasa los 3700 m, en sus límites altitudinales más altos. Es aquí donde existe una dinámica alta de deslizamientos, los mismos que si bien afectan la diversidad vegetal, también son considerados un motor generador de continuos procesos de regeneración. Se establecieron 138 parcelas, distribuidas a o largo de un gradiente latitudinal y altitudinal, de 1 x 5m al occidente del PNP, encontrándose 218 especies representantes de 180 géneros y 51 familias; el piso altitudinal de mayor diversidad está a los 2700 m. El análisis TWINSPAN separó nueve comunidades vegetales, mientras que el análisis multivariado CANOCO, determinó que las características edafológicas influyen en la composición de la vegetación, mientras que la altitud y la pendiente no establecen notable diferencia.
Palabras clave. Deslizamientos naturales, flora pionera, gradiente altitudinal, Parque Nacional Podocarpus-Ecuador.
ABSTRACT
Podocarpus National Park in Southern Ecuador occupies a climatological key position position with regard to the Huancabamba Depression, which forms a natural biogeographical barrier in the South. The dissected geography of this area, in addition to a thin vegetation cover and high annual rainfall, exceeding 6000mm in the Paramo areas, lead to severe landslide effects on the western and eastern flanks of the Cordillera Oriental, which reaches less than 3700 m at its highest point. The landslide dynamics in the region are the driving force generating constant regeneration processes in wide areas. This study established 138 plots of 1x5m on the western side of Podocarpus National Park. Twohundred-eighteen plant species in 180 genera and 51 families were registered, with the highest diversity found at 2700 m. The TWINSPAN analysis separated the plots into nine vegetation communities, while multivariate CANOCO analysis delineated soil characteristics as an important distinguishing factor for vegetation composition. Factors like slope and altitude did not have a remarkable influence on the vegetation communities.
Key words. Naturals landslides, pioneer flora, altitudinal gradient, Podocarpus National Park-Ecuador.
INTRODUCCIÓNEcuador es reconocido como un país megadiverso, el registro de plantas vasculares señala 17058 especies (Ulloa Ulloa & Neill 1999-2004), de las cuales el 26 % de la flora nativa es endémica (Valencia et al. 2000). Los Andes al sur del Ecuador no tienen una clara dirección Norte Sur, sino que se bifurcan y descienden a sus límites altitudinales más bajos, denominada la "deflexión de Huancabamba", área de ubicación del PNP en las provincias de Loja y Zamora Chinchipe. Los bosques montanos y páramos del Podocarpus son considerados "perhúmedos", ya que se han registrado precipitaciones sobre los 6000 mm (Richter 2003), las cuales provienen principalmente de la región Amazónica, debido a las convergencias intertropicales de masas de aire con promedios de humedad de 75 a 80% (Luteyn 1999), condiciones aptas para un alto epifitismo (Bussmann 2001) y endemismo (Lozano et al. 2003), mientras que más al sur las condiciones se vuelven más secas (Luteyn 1999).
]]> Los estudios de sucesión vegetal desempeñan un importante papel en la ecología de plantas (Peet & Christensen 1980); éstos poseen una larga trayectoria de investigación, tratando de entender los diferentes procesos que los conllevan en su regeneración natural (White & Jentsch 2001). Los daños causados por los deslizamientos en la naturaleza, aun que pueden llegar a ser tanto desastrosos como muy importantes, han sido poco estudiados en los Andes del Ecuador y se desconocen varios aspectos en su dinámica de sucesión (Stern 1995, Ohl & Bussmann 2004). Estos procesos están influenciados por una serie de factores como pendiente, tipo de suelos, temblores. Ohl & Bussmann (2004) señalan que la frecuencia de deslizamientos naturales en el PNP es muy elevada debido a condiciones extremas como: lluvias largas e intensas que provocan la inestabilidad de los suelos, fuertes pendientes, poca profundidad de las raíces de las plantas, sumado a la litología del lugar. Estos factores inducen los procesos de deslizamiento, que además de ser un problema geológico, influyen en cambios significativos sobre la vegetación y la regeneración natural de hábitats frágiles.El presente estudio se orienta a conocer la dinámica de la regeneración y variación vegetal en la gradiente altitudinal y latitudinal en diferentes sitios del sector Occidental del PNP, a través de la relación de los factores ambientales, la flora pionera y los grupos taxonómicos importantes que se presentan para cada piso ecológico, determinando su influencia en la recuperación y la dinámica.
Zonas de vida del Parque Nacional Podocarpus
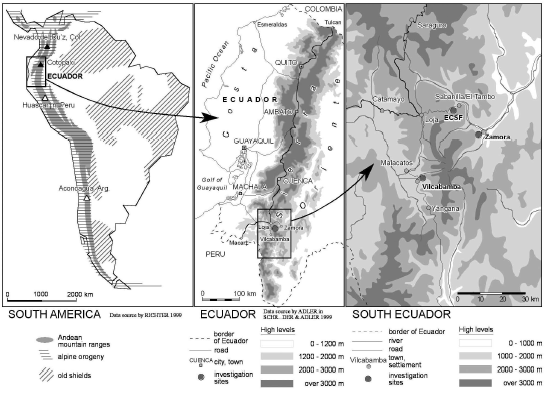
El PNP (Fig. 1) fue creado en 1982 (Apolo 1984), posee una superficie de 146 280 ha, entre los rangos altitudinales 1000 a 3600 m. Debido a que el Parque se extiende en dos regiones diferentes (andina y amazónica), según Holdridge (1978), posee seis zonas de vida:
1) bosque húmedo pre-montano.- Se lo encuentra ubicado en el sector sur "Yangana", bajo los 2000 m.s.n.m, con una temperatura promedio de 18ºC.
2) bosque muy húmedo pre-montano.- Esta zona se halla comprendida entre 1000 y 2000 m.s.n.m, la temperatura varía entre 18ºC y 24ºC. Constituye la zona más baja del Parque Nacional Podocarpus, pues pertenece a las cuencas de los ríos Nangaritza, Chumbiratza y Bombuscaro. En esta zona, la vegetación es exuberante, propia de la Amazonía.
3) Bosque muy húmedo montano-bajo.- Representa la mayor área del Parque y se sitúa entre 2000 y 3000 m.s.n.m, con una temperatura variable entre 12 y 18ºC. Una buena parte de esta zona asciende por las estribaciones de la cordillera Real, donde la vegetación disminuye en altura, formando un bosque que se pierde en la neblina hasta alcanzar los 2800 m.s.n.m "Ecotono" y luego se transforma en un paisaje de páramo. La parte más baja de esta zona es el hábitat óptimo del "romerillo" Podocarpus oleifolius, árbol que ha dado su nombre al Parque; la mayor densidad de población de esta especie se localiza en las cuencas de los ríos Numbala y Loyola. En esta formación se desarrollaron las investigaciones del presente estudio. ]]>
Área de estudio
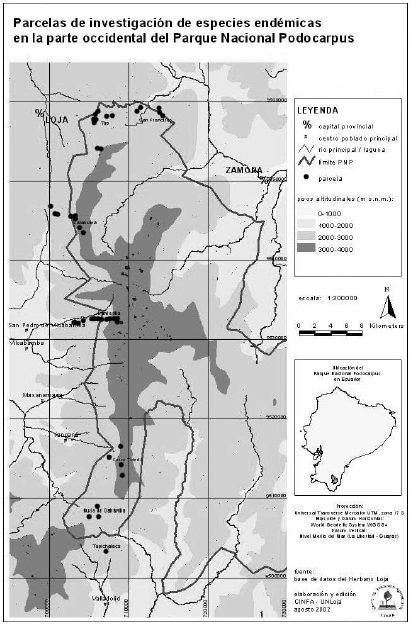
La zona de estudio se ubica en la franja occidental, en el norte desde el sector El Tiro, hasta Sabanilla en el sur. Abarca una extensión de 17 435 ha, su altitud va de 2100 a 3300 m.s.n.m. Las zonas involucradas en la investigación se ubican bajo las siguientes coordedas U.T.M.:
Norte: Sector El Tiro, rango altitudinal entre 2700 a 3200 m, coordenadas 9555054,13 de latitud Norte y 705702,45 de longitud Este.
Centro Norte: Sector Cajanuma, rango altitudial entre 2500 hasta 3200 m, coordenadas 9559676,87 de latitud Norte y 701760,25 de longitud Este.
Centro Sur: Sector Vilcabamba, rango altitudinal entre 2000 a 4000 m, coordenadas 9532056,17 de latitud Norte y 703239,58 de longitud Este.
Sur: Sector Cerro Toledo, rango altitudinal entre 2950 a 3400 m, coordenadas 9515049.98 de latitud Norte y 710414.42 de longitud Este.
Sur: Sector cordillera de Sabanilla, rango altitudinal entre 2700 a 2900 m, coordenadas 9508701,62 de latitud Sur y 706081,47 de longitud Este. En la figura 2, se presenta la ubicación de las parcelas de norte a sur.
MATERIALES Y MÉTODOS
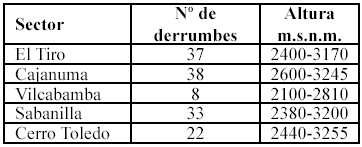
El área de estudio se dividió en cinco sectores (Tabla 1), en tres rangos altitudinales que abarcaban de 2100-2600 de 2600-3000 y de 3000-3400 m.s.n.m. Dentro de cada rango se estimó un número similar de deslizamientos.
Para el estudio de los parámetros ecológicos en los deslizamientos se usó la metodología de Braun-Blanquet (1979), se instalaron unidades de muestreo consistentes en parcelas de 1x 5 m, las cuales se subdividieron en cinco sub-parcelas de 1 m2, donde se identificaron y contabilizaron cobertura en porcentajes de briofitas en general (incluyen líquenes y musgos) sin identificación de generos, especies del estrato herbáceo y arbustivo en porcentajes, se tomó en cuenta tres rangos de altura de las especies: 0-15; de 15-30 y > 30 cm. En algunos de los casos, se establecieron de dos a tres parcelas en un mismo deslizamiento, considerando una distancia de 10 metros entre cada parcela (Figura 3).

El material herborizado se identificó en el Herbario de la Universidad Nacional de Loja (LOJA), para la identificación del material se uso Harling & Andersson (1974-2003), y la nomenclatura estuvo basada en Brako & Zarucchi (1993) y Jørgensen & León-Yánez (1999). Las colecciones se compararon en el Herbario de la Pontificia Universidad Católica (QCA). Ejemplares de todas las especies se encuentran depositadas en el Herbario LOJA.
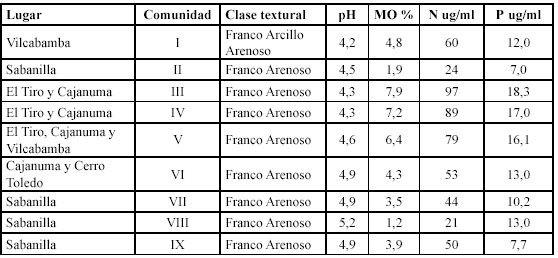
La caracterización de los suelos se realizó mediante barrenaciones hasta una profundidad de 30 cm. Se realizaron seis muestreos por sector, distribuidos a lo largo de la franja norte-sur y en el gradiente altitudinal. Los análisis físico-químicos, textura, materia orgánica (MO), acidez (pH), fósforo (P) y nitrógeno disponible (N), se realizaron en el Laboratorio de Suelos de la Universidad Nacional de Loja.
Agrupación de Comunidades Vegetales
Para realizar la agrupación de los datos de las especies en comunidades, se utilizó el programa TWINSPAN (Hill 1994), el cual presenta una matriz de ordenamiento de dos entradas, la primera son las parcelas y las especies como variable, llenándose con presencia o ausencia (1 o 0) de especies en una tabla excel, la misma que es transportada al programa, al correr o ejecutarse el programa, se agrupan o separan las diversas especies en comunidades, con base en la mayor similitud en la composición florística, la cual está separada por un "eigenvalor" entre (0-1).
Los "eigenvalores" que se muestran en el dendrograma, indican el grado de separación. Los valores superiores a 0,5 señalan que la separación es fuerte porque la composición florística entre comunidades es muy diferente, mientras que los valores inferiores a 0,2 indican que la composición florística entre comunidades es similar. El programa además separa los grupos fitosociológicos, en "Especies Exclusivas", que son aquellas especies que solamente ocurren en una sola comunidad, es decir que no están presentes en el resto de comunidades. "Especies Compartidas", especies presentes en dos o más comunidades y tienen una alta representatividad entre las comunidades. "Especies Generalistas", especies que ocurren en todas las comunidades.
Análisis entre Factores Edáfico-Ambientales y la Vegetación con el Uso de CANOCO
Con los datos de suelos de las barrenaciones realizadas en campo y con los datos registrados en el campo de posición geográfica, altitud, inclinación y exposición, se aplicó el programa de Análisis de Correspondencia Canónica "CANOCO" (Ter Braak 1998), el cual se basa en la regresión múltiple de ordenación de muestras, los valores de las variables y los factores ambientales se representan en un espacio imaginario con cuatro ejes. Este programa usa los datos que se derivan de TWINSPAN y sobre ellos aporta que factores ambientales son la respuesta de cada tipo de comunidad vegetal.
Grupos Taxonómicos Importantes y Característicos Presentes en los Deslizamientos
Para la identificación de los grupos taxonómicos importantes y característicos que ocurren en los deslizamientos, se tomó en cuenta los datos de las nueve comunidades obtenidas por el programa TWINSPAN, con los cuales se procedió a la determinación de parámetros ecológicos de densidad, (Número total de indiv./total de área muestreada); densidad relativa (Número de individuos por especie/número total de especies x 100); diversidad relativa (Número de especies/número total de especies x 100) por familia. ]]>
Definición de cambios florísticos de la regeneración en la gradiente altitudinalSe realizó la diferenciación florística en la gradiente altitudinal mediante el índice de similitud de Sorensen que consideró la comparación de las especies comunes existente, entre las nueve comunidades se establece la semejanza en la composición florística entre los sitios. Además se aplicó el índice de diversidad de Simpson para comparar sitios con base en la diversidad.
Índice de Similitud de Sorensen:
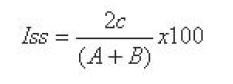
Donde:
Iss = Índice de Similitud de Sorensen.
A = Número de especies de la muestra 1
B = Número de especies de la muestra 2
c = Número de especies en común.
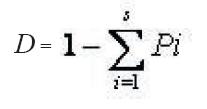
D =
Donde:
D = Índice de diversidad de Simpson
Pi = Proporción de individuos de la especie i en el sitio
RESULTADOS
En la zona estudiada se identificaron 218 especies, dentro de 180 géneros en 51 familias. Resultados que verifican un alto grado de regeneración natural. Ohl & Bussmann (2004) reportan en su investigación 40 familias botánicas representadas por 32 géneros y 146 especies vegetales presentes en deslizamientos naturales del sector de San Francisco parte oriental del PNP.
Los grupos taxonómicos más importantes están representados por las familias Asteraceae con 34 especies en 19 géneros, Melastomataceae con 19 especies en siete géneros y ERICACEAE con 18 especies en nueve géneros. De estos grupos las especies pioneras más importantes en densidad son: Ericaceae (Disterigma alaternoides Gaultheria erecta, Bejaria aestuans, Gaultheria reticulata, Gaultheria strigosa, Vaccinium floribundum); Melastomataceae (Brachyotum benthamianum, Brachyotum rugosum, Miconia dodsonii, M. subulata); Asteraceae (Ageratina dendroides, A. cutervensis, Baccharis genistelloides, B. obtusifolia, Gynoxys buxifolia); Poaceae (Cortaderia bifida). Según los grupos fitosociológicos, se destacan los siguientes:
]]> Grupo de especies exclusivasGrupo de especies compartidas
1). Subgrupo de Rinchospora vulcani, Weinmannia glabra y Brachyotum campii, para las comunidades VI y VII.
2). Subgrupo de Campyloneurum sp., y Elleanthus aurantiacus, para la comunidad VIII y IX.
Grupo de especies generalistas
Las especies generalistas son Sticherus revolutus, Disterigma alaternoides, Gaultheria erecta, Cortaderia bifida, Gaultheria reticulata, Blechnum cordatum, Lycopodium complanatum y Vaccinium floribundum.
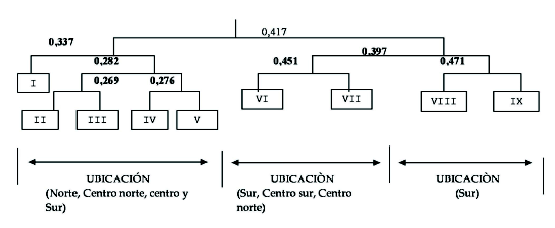
El análisis TWINSPAN de los 138 levantamientos ordena las comunidades vegetales como se muestra en la figura 4, en la que define los niveles de separación de la zona de estudio en nueve comunidades vegetales.
]]>
Figura 4. Representación gráfica de las comunidades vegetales.Se definen tres sectores la primera: norte, centro norte y centro sur (I, II, III, IV y V); la segunda: sur, centro sur y centro norte (VI y VII); la tercera: sur (VIII y IX). La descripción de cada comunidad sucesional se detalla a continuación:
Comunidad Sucesional I. Baccharis genistelloides y Tibouchina laxa (2100 m)
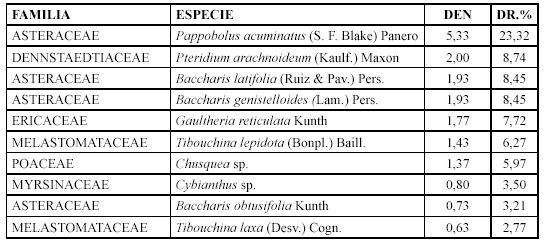
Se ubica en el sector centro Occidental del PNP, parte alta de Vilcabamba, en esta comunidad se agruparon tres parcelas a una altitud de 2100 m, con una inclinación de 50 % la que se define como pendiente escarpada. Se registraron 38 especies dentro de 34 géneros en 20 familias. Las especies que caracterizan esta comunidad son: Baccharis genistelloides, Tibouchina laxa, Gaultheria reticulata, Cronquistianthus niveus y Weinmannia fagaroides.
Las especies con mayor densidad relativa son: Pappobolus acuminatus, Pteridium arachnoidium y Baccharis latifolia (Tabla 2). Mientras que los géneros con mayor diversidad relativa son: Baccharis con tres especies (8,82 %), Rhynchospora con dos (5,88 %) y Tibouchina con dos (5,88 %).
Comunidad Sucesional II. Gaultheria erecta y Brachyotum bentamianum (2430 m)
Se encuentra ubicada en el Sur del sector de Sabanilla, tres parcelas determinaron esta comunidad, en la cota altitudinal de 2430 m, con una inclinación entre 50 - 55 % la cual se denomina como pendiente escarpada. Se registraron 31 especies dentro de 24 géneros en 12 familias. Las especies que caracterizan esta comunidad son: Ageratina dendroides, Baccharis genistelloides, Brachyotum benthamianum, Pteridium arachnoideum, Disterigma alaternoides y Gaultheria erecta.
]]> Las especies con mayor densidad relativa son: Gaultheria erecta, Sticherus revolutus y Disterigma alaternoides (Tabla 3). Mientras que los géneros con mayor diversidad relativa son Baccharis con tres especies (12,50 %), Disterigma y Vaccinium con dos (8,33).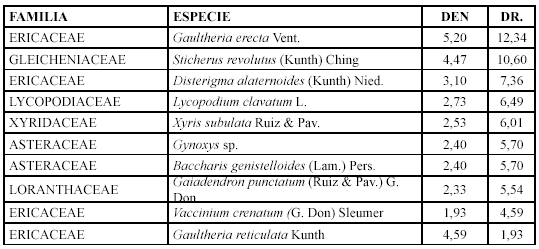
Comunidad Sucesional III. Stipa ichu y Paepalanthus ensifolius (2400 - 2850 m)
Se encuentra ubicada en el norte y centro del sector Occidental del PNP y, en El Tiro y Vilcabamba, cuenta con 34 parcelas en un rango altitudinal entre 2400 - 2850 m.s.n.m y con una inclinación entre 40 - 70 %, predominando las pendientes muy escarpadas. Se registraron 64 especies dentro de 51 géneros en 27 familias. Las especies que caracterizan esta comunidad son: Ageratina dendroides, Baccharis genistelloides, Brachyotum benthamianum, Sticherus revolutus, Orthrosanthus chimboracensis, Paepalanthus ensifolius, Stipa ichu, Blechnum cordatum y Gaultheria erecta.
Las especies con mayor densidad relativa son: Sticherus revolutus, Stipa ichu y Baccharis genistelloides (Tabla 4).
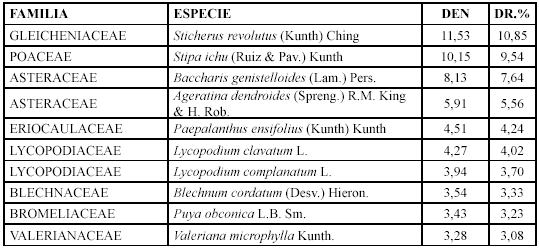
Comunidad Sucesional IV. Sticherus revolutus y Loricaria thuyoides (2750-3235 m)
Se encuentra ubicada en el Norte y centro Norte del sector Occidental del PNP, en El Tiro y Cajanuma respectivamente, cuenta con 25 parcelas en un rango altitudinal entre 2750 - 3235 m.s.n.m., y con una inclinación entre 42 - 72 % predominando las pendientes muy escarpadas. Se registraron 49 especies dentro de 37 géneros en 24 familias. Las especies que caracterizan esta comunidad son: Bejaria aestuans, Freziera minima, Sticherus revolutus, Loricaria thujoides, Disterigma alaternoides y Cortaderia bifida.
Las especies con mayor densidad relativa son: Sticherus revolutus, Loricaria thujoides y Freziera minima (Tabla 5).
Mientras que los géneros con mayor diversidad relativa son: Brachyotum con tres especies (8,10 %), Bejaria y Disterigma con dos (5,40 %).
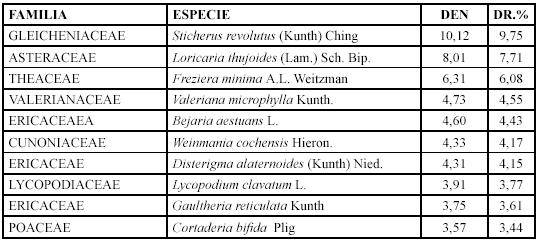
Comunidad Sucesional V. Gaultheria erecta e Hypericum lancioides (2650 - 3170 m)
Se encuentra ubicada en el norte, centro Norte y centro de la parte Occidental del PNP y, pertenece al sector de El Tiro, Cajanuma y Vilcabamba respectivamente, cuenta con 20 parcelas en un rango altitudinal entre 2760 - 3170 m.s.n.m., y con una inclinación entre 38 - 74 %, siendo características las pendientes muy escarpadas. Se registraron 54 especies dentro de 42 géneros en 25 familias. Las especies que caracterizan esta comunidad son: Gaultheria erecta, Hypericum lancioides, Disterigma alaternoides, Blechnum cordatum, Arcytophyllum setosum, Bejaria aestuans, Sticherus revolutus, Lycopodium clavatum, Disterigma empetrifolium, Gaiadendron punctatum y Orthrosanthus chimboracensis.
]]> Las especies con mayor densidad y densidad relativa son: Hypericum lancioides, Bejaria aestuans y Gaultheria erecta (Tabla 6).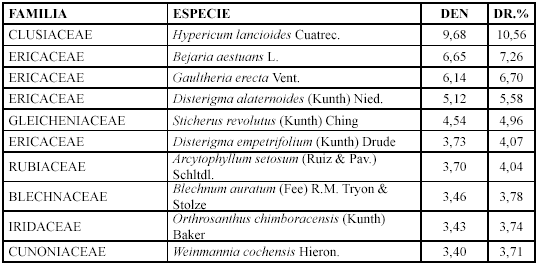
Mientras que los géneros con mayor diversidad relativa son: Weinmannia con cuatro especies (9,52 %), Brachyotum y Blechnum con tres (7,14 %).
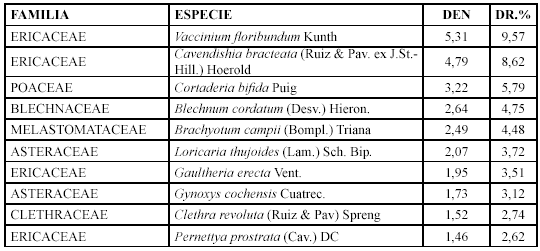
Comunidad Sucesional VI. Blechnum cordatum y Vaccinium floribundum (2440 - 3200 m)
Esta comunidad se encuentra ubicada en el centro Norte y centro sur del sector Occidental del PNP, en los sitios de Cajanuma y Cerro Toledo, en esta comunidad se agruparon 23 parcelas, de los cuales una pertenece al sector de Cajanuma que se encuentra a una altitud de 3080 m.s.n.m, mientras que las de Cerro Toledo se encuentran en un rango altitudinal de 2440 a 3200 m.s.n.m., y con una inclinación entre 40 - 72 %, siendo características las pendientes muy escarpadas. Se registraron 63 especies dentro de 45 géneros en 25 familias. Las especies que caracterizan esta comunidad son: Blechnum cordatum, Cortaderia bìfida, Vaccinium floribundum, Brachyotum campii, Cavendishia bracteata, Gnaphalium elegans y Gynoxys cuicochensis.
Las especies con mayor densidad relativa son: Vaccinium floribundum, Cavendishia bracteata y Cortaderia bifida. (Tabla 7).
]]>
Tabla 7. Densidad y densidad relativa de las especies de la Comunidad VI.Mientras que los géneros con mayor diversidad relativa son: Brachyotum con cinco especies (11,11 %), Gaultheria y Gynoxys con tres (6,66 %).
Comunidad Sucesional VII. Elaphoglosum lingua y Sticherus revolutus (2850-3190 m)
Se encuentra ubicada al Sur del PNP y, pertenece al sector de Sabanilla exclusivamente, cuenta con 14 parcelas en un rango altitudinal de 2850 - 3190 m.s.n.m., y con una inclinación entre 55 - 75 % siendo estas pendientes muy escarpadas. Se registraron 55 especies dentro de 44 géneros en 23 familias. Las especies que caracterizan esta comunidad son: Elaphoglosum lingua, Sticherus revolutus, Blechnum auratum, Disterigma alaternoides, Oxalis spiralis y Cortaderia bìfida.
Las especies con mayor densidad y densidad relativa son: Disterigma alaternoides, Sticherus revolutus y Oligactis coriacea (Tabla 8).
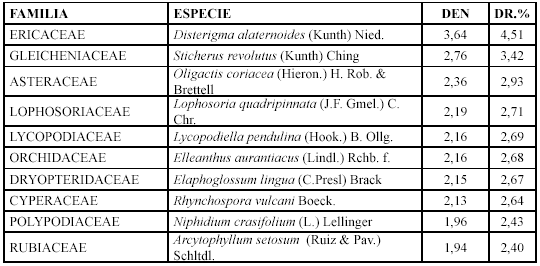
Mientras que los géneros con mayor diversidad relativa son: Blechnum con tres especies (6,81 %), Gaultheria y Disterigma con dos (4,54 %).
Comunidad Sucesional VIII. Ellanthus aurantiacus y Disterigma alaternoides (2580-2610 m)
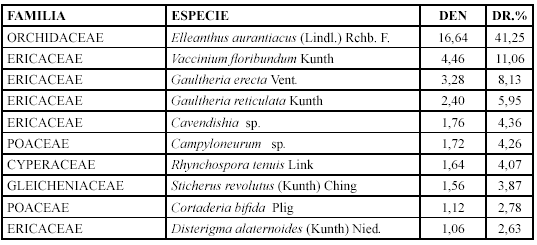
Mientras que los géneros con mayor diversidad relativa son: Gaultheria con tres especies (17,64 %), Vaccinium y Lycopodium con dos (11,76 %).
Comunidad Sucesional IX. Disterigma alaternoides y Gaultheria erecta (2380 - 3080 m)
Se encuentra ubicada al Sur del PNP en el sector de Sabanilla, cuenta con 11 parcelas en un rango altitudinal de 2380 - 3080 m.s.n.m., con una inclinación entre 55 - 78 %, siendo estas pendientes muy escarpadas. Se registraron 24 especies dentro de 21 géneros en 13 familias. Las especies que caracterizan esta comunidad son: Disterigma alaternoides, Gaultheria erecta, Cortaderia bìfida, Elleanthus aurantiacus, Pitcairnia pungens y Baccharis latifolia.
Las especies con mayor densidad y densidad relativa son Gaultheria reticulata, Gaultheria foliolosa y Elleanthus aurantiacus (Tabla 10).
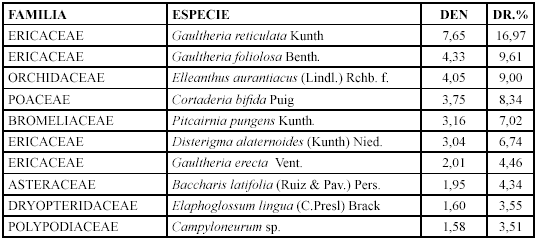
Mientras que los géneros con mayor diversidad relativa son: Gaultheria con tres especies (14,28 %), Disterigma y Pernettya con una (4,76 %).
Índice de similitud de Sorensen
Los resultados obtenidos (Tabla 11), indican que las comunidades IV y V presentan mayor similitud en la composición florística con un valor de 61, se asume que este resultado esta apoyado por la cercanía de los sitios El Tiro (Norte) y Cajanuma (centro Norte).
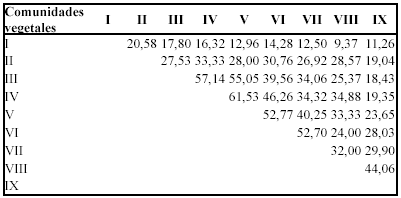
La pendiente en estas comunidades es similar debido a que se encuentran entre rangos de 38 - 70 %, predominando las pendientes muy escarpadas.
El segundo valor más elevado de similitud en la composición florística corresponde a las comunidades III y IV, lo que también se puede explicar debido a lo cercano de los sitios a los que éstas pertenecen Vilcabamba (centro sur) y Cajanuma.(centro sur). Además existe similitud en factores como: pendiente en la cual tienen rangos muy elevados que van desde los 60 - 75 %, por lo que se considera como muy escarpadas, así mismo comparten rangos altitudinales muy similares que abarcan desde los 2400 a 3200 m.
]]> Las comunidades I y VIII son las que menor similitud presentan, debido a que se encuentran distanciados y a que la comunidad I se encuentra únicamente en el sector parte alta de Vilcabamba a una altitud de 2100 m.s.n.m (sector centro de la zona de estudio), mientras que la comunidad VIII está ubicada exclusivamente en el sector de Sabanilla con una altitud mayor a 2 400 m.s.n.m (sector sur de la zona de estudio).Índice de diversidad de Simpson (tabla 12)

Las comunidades con mayor diversidad son las comunidades III y IV, la comunidad III cuenta con 107 especies dentro de 41 familias, ésta se encuentra ubicada en los sectores de El Tiro y Cajanuma (norte y centro), La comunidad IV se encuentra ubicada en el sector de Sabanilla, cuenta con 60 especies dentro de 29 familias.
Diversidad de especies en el gradiente
La distribución del número de especies en el gradiente altitudinal, se determino con base en la altitud por número de especies (Figura 5).
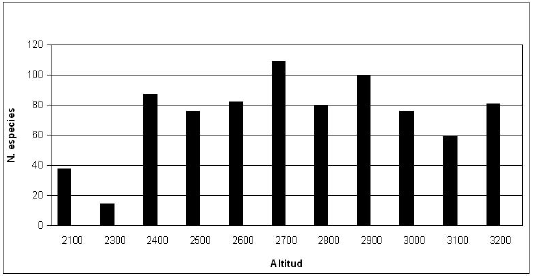
]]>
Figura 5. Distribución del número de especies en el gradiente altitudinal.De las 218 especies registradas, se evidencia que la mayor diversidad de especies en la regeneración natural se localiza entre un rango altitudinal que va desde 2400 m. con un número de 90 especies, a los 2900 m. se concentran 101 especies y a los 2700 m, existe la mayor agrupación con 106 especies. Determinando que existe cambios en la composición florística y que hay mayor regeneración y diversidad de especies en el rango altitudinal de 2700 m, franja que pertenece al bosque montano alto, estudios comparativos Lozano et al. (2003), menciona que el rango altitudinal donde se concentran las especies endémicas está entre 2700 a 3400 m.s.n.m, con mayor concentración en 2800 a 3200 m.s.n.m.
Suelos de los sitios de estudio
Los suelos de los sitios estudiados son fuertemente ácidos, con escasa presencia de materia orgánica, pobres en nitrógeno y fósforo aprovechable, los suelos sufrieron la pérdida de horizontes, predominando el afloramiento de roca, sin embargo la regeneración natural, se explica, según lo señalan Teuscher & Adler (1985), gran mayoría de plantas que prosperan en suelos ácidos, no solamente toleran estas condiciones, sino que efectivamente las necesitan, porque sus procesos metabólicos están adaptados a ellas y dependen de valores bajos del pH, por tal motivo son incapaces de obtener sus nutrimentos de un suelo alcalino o de un suelo neutro, habiendo sido comprobado por las investigaciones fisiológicas. En la tabla 13, se presenta los resultados para cada comunidad.
Estos suelos se caracterizan por ser moderadamente pesados a livianos es decir de textura franco a franco arenosos, estas texturas se caracterizan por tener buena capacidad de retención de agua, no son compactos lo cual permite el fácil movimiento de agua y aire, lo que da facilidad para la penetración de raíces y disponibilidad de los elementos nutritivos (Lozano et al. 2003), tomando estos referentes se puede entender la posibilidad de la regeneración natural de especies vegetales en los deslizamientos naturales.
La hoja geológica Gonzanama (hoja 57, escala 1:100 000), determino que los sectores en donde se realizó los muestreos son de origen de rocas metamórficas, de la serie Zamora. Las rocas de este sector consisten principalmente de filitas, cuarcitas y mico esquistos; las cuarcitas predominan al Este y al Oeste de Loja los esquistos y gneises biotiticos, conjuntamente con esquistos grafiticos. El rumbo de foliación es generalmente NNE (Nor - noreste), con ángulos de 55 a 78 º.
El análisis de los factores ambientales representado en la figura 6, indica que las comunidades I y VI se encuentran directamente influenciadas por el pH, pendiente, siendo estos suelos franco arcillo arenosos y, en menor grado influenciadas por el potasio (K) y alturas que van desde los 2 100 a 3 200 m, pues son variables que se encuentran más cerca del eje central y en la misma dirección. Además se encuentran inversamente influenciadas por el fósforo (P) aprovechable.
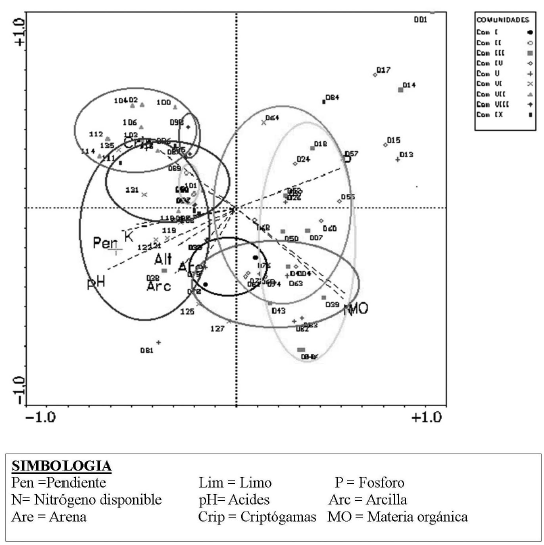
Las comunidades II, VII, VIII y IX, poseen suelos franco arenosos con gran cantidad de limo y baja cantidad de materia orgánica, están inversamente influenciadas por materia orgánica y nitrógeno, poseen pendientes muy escarpadas que sobrepasan el 55 %. Mientras que las comunidades III, IV y V están determinadas por suelos franco arenosos y gran cantidad de materia orgánica (MO), se encuentran influenciadas directamente por Fósforo (P), Nitrógeno (N) aprovechable.
Los datos de Ohl & Bussmann (2004), en el análisis físico-químico de los suelos de los deslizamientos en el sector oriental del PNP (Estación San Francisco), no muestra diferencias mayores entre los sitios muestreados. Y no existe ninguna correlación fuerte entre la distribución de vegetación y características de los suelos. No obstante se anota que los deslizamientos que se encuentran en altitudes más bajas tienden a tener una textura más arenosa.
CONCLUSIONES
Dentro de la zona de estudio, que abarca los sectores: El Tiro, Cajanuma, Vilcabamba, Cerro Toledo y Sabanilla, se registraron IX comunidades vegetales y los "eingenvalores" obtenidos en el dendograma, en su primera separación indican que la diferencia florística que existe entre las comunidades ubicadas en el norte, centro-norte y centro, de las ubicadas en el centro-sur y sur, no es fuerte y por lo tanto la composición florística en su mayoría es similar entre cada Sector.
La mayor similitud de especies existe entre la comunidad IV y V, las cuales se encuentran ubicadas en los sectores de El Tiro y Cajanuma.
]]> Los grupos taxonómicos más importantes se encuentra representados por las siguientes familias: Asteraceae con 34 especies en 19 géneros, Melastomataceae con 19 especies en 7 géneros y Ericaceae con 18 especies en 9 géneros. De estos grupos las especies pioneras más importantes en densidad son: Disterigma alaternoides Gaultheria erecta, Bejaria aestuans, Gaultheria reticulata, Gaultheria strigosa, Vaccinium floribundum (Ericaceae), Brachyotum benthamianum, Brachyotum rugosum (Melastomataceae), Ageratina con 3 especies: Ageratina dendroides, Baccharis genistelloides, (Asteraceae), Cortaderia bifida (Poaceae).Se evidencia que la mayor diversidad de especies en la regeneración natural se localizan entre un rango altitudinal que va desde 2400 m.s.n.m. con un número de 90 especies, luego a 2900 m.s.n.m. 101 especies y finalmente a 2700 m.s.n.m. 109 especies, siendo este el más representativo, rangos que se encuentran principalmente en la franja de bosque montano alto.
Lozano et al. (2003), anotan que los factores ambientales y edáficos no tienen relación significativa con el tipo de vegetación o tipos de comunidad. También Volgelegt y Contanze (2000), sugieren que no existe ninguna relación fuerte entre la distribución de vegetación y características de la tierra, sin embargo los cambios en los factores climáticos debido a la altitud (aumento de humedad, despliegue de temperatura, insolación creciente y vientos más fuertes) es muy importante para la composición de la vegetación. Lo que se corrobora en esta investigación, ya que las especies encontradas son comunes en los diferentes sitios de estudio y existe una escasa diferencia en la composición florística.
Se define la franja altitudinal entre los 2700 a 2800 como la que mayor aporte de recursos genéticos otorga a los deslizamientos naturales, coincide con un análisis de endemismo y diversidad realizado por Lozano et al. (2003) donde señala que es a los 2800 m, franja denominada "ecotono", el cual es la transición de bosque nublado a páramo, donde se concentra la mayor diversidad y endemismo al occidente del PNP. Por otro lado la mayor diversidad se concentró en el sector de Cajanuma y El Tiro al centro-norte y norte del parque. La presencia de abundancia de especies pertenecientes a las familias Asteraceae; Melastomataceae y Ericaceae, nos indican que en el caso de las dos primeras existe una rápida recolonización debido a la producción de semillas y dispersión por anemofilia que facilita su movilización, en el caso de las Ericaceas algunas Melastomataceas se asume que sus drupas son fácilmente dispersadas y escarificadas por aves, con un alto porcentaje de germinación en suelos jóvenes con poca materia orgánica.
AGRADECIMIENTOS
Los autores desean dejar constancia de agradecimiento al Instituto de Botánica de la Universidad de Hohenheim, Stuttgart-Alemania y al proyecto DFG FOR 402-1/1 TP7. A Bolívar Merino curador del herbario LOJA, a Zhofre Aguirre Director del herbario LOJA, a Paola Sarango, Anja Meineke, Wendy Warries, Lea Goepfert quienes apoyaron esta investigación. A las Fundaciones JOCOTOCO y Naturaleza y Cultura Internacional.
LITERATURA CITADA
1. APOLO, W. 1984. Plan de Manejo del Parque Nacional Podocarpus. Ministerio de Agricultura y Ganadería, Quito. [ Links ]
2. BUSSMANN, R. 2001. Epiphyte Diversity in a Tropical Andean Forest-Reserva Biológica San Francisco, Zamora-Chinchipe, Ecuador. Ecotropica 7: 43-59. [ Links ]
3. BRAKO, L. & J. L. ZARUCCHI. 1993. Catalogue of the flowering plants and Gymnosperms of Peru. Monographs in Systematic Botany 45. [ Links ]
4. BRAUN BLANQUET, J. 1979. Fito Sociología-Bases para el Estudio de las Comunidades Vegetales. España. [ Links ]
5. HARLING, G. & L. ANDERSSON (eds.). 1986-2003. Flora of Ecuador 25-60. [ Links ]
6. JØRGENSEN, P.M. & S. LEÓN-YÁNEZ. 1999. Catalogue of the Vascular Plants of Ecuador. Monographs in Systematic Botany 75: 1-1181. [ Links ]
7. HILL, M.O. 1994. DECORANA and TWISPAN, for ordination and classification of multivariate species data: a new edition, together with supporting programs, in FORTRAN 77. Huntingdon. Pp. 57. [ Links ]
8. HOLDRIDGE, L. 1978. Ecología: basadas en las zonas de vida. Trad. Humberto Jiménez Saa. San José, C. R., Instituto Interamericano de Ciencias Agrícolas. Pp. 216 [ Links ]
9. LOZANO, P., T. DELGADO & Z.M. AGUIRRE. 2003. Estado Actual de la Flora Endémica Exclusiva y su Distribución en el Occidente del Parque Nacional Podocarpus. Publicaciones de la Fundación Ecuatoriana para la Investigación y Desarrollo de la Botánica. Loja, Ecuador. Pp. 180. [ Links ]
10. OHL, C. & R.W. BUSSMANN. 2004. Recolonisation of Natural Landslides in Tropical Mountains Forest of Southern Ecuador. Feddes Repertorium 115: 3-4; 248-264. [ Links ]
11. LUTEYN, J. 1999. Páramos a checklist of plant diversitygeographical distribution, and botanical literature. Memoirs of The New York Botanical Garden 84: 1- 278. [ Links ]
12. PEET, R.K. & L. CHRISTENSEN. 1980. Succession: A Population Process. Vegetation 43: 131-140. [ Links ]
13. RICHTER, M. 2003. Using Epiphytes and Soil Temperatures for Eco-Climatic Interpretation in Southern Ecuador. Erdkunde 57: 161-181. [ Links ]
14. STERN, M.J. 1995. Ecosystem Response to Natural and Anthropogenic Disturbances in the Andean Cloud Forest of Ecuador. Pp. 146. Ph.D. Thesis. University of California. [ Links ]
15. TER BRAAK, C.J. 1988. Canoco: A Fortran program for canonical community ordination. Microcomputer Power, Ithaca, New Cork. Pp. 299. [ Links ]
16. TEUSCHER, H. & R. ADLER. 1985. El Suelo y su fertilidad. Mexico. CECSA. 9. Pp. 132. [ Links ]
17. ULLOA ULLOA, C. & D. NEILL. 1999-2004. Cinco años de adiciones a la Flora de Ecuador 1999-2004. Universidad Técnica particular de Loja. [ Links ]
18. VALENCIA, R., N. PITMAN, S. LEÓN-YÁNEZ & P.M. JØRGENSEN (eds) 2000. Libro rojo de las plantas endémicas del Ecuador 2000. Herbario QCA, Pontificia Universidad Católica del Ecuador, Quito, Ecuador. Pp. 1-489. [ Links ]
19. VOLGELEGT, V. & C. OHL. 2000. Vegetation on natural landslides in the tropical montane forest of souther Ecuador with special consideration of the altitudinal gradient. Diplomarbeit zur Erlansung des grades einer Diplom - Biogeographin. Saarbruchen. Universitat des Saarlandes. Pp. 14-69. [ Links ]
20. WHITE, P. & A. JENTSCH. 2001. The Search for Generality in Studies of Disturbance and Ecosystem Dynamics. Progress in Botany 62: 399-449. [ Links ]
Recibido: 05/02/2007
Aceptado: 06/02/2008