Effect of variation of body weight and body condition and leptin and luteinizing hormone receptor expression, on postpartum anovulation of zebu (Bos indicus) cows.
Efeito das variações de peso e condição corporal, de lá expressão de receptores de leptina e hormôna luteinizante, na anovulação pós parto em vacas zebu (Bos indicus).
Carlos A Giraldo1*, MV, MS; Martha Olivera1, MV, Dr Sci Agr; Zulma T Ruíz Cortés1, MV, PhD.
1Grupo Reproducción – Fisiología y Biotecnología, Facultad de Ciencias Agrarias, Universidad de Antioquia, AA 1226, Medellín, Colombia.
]]> (Recibido: 6 junio, 2007; aceptado: 30 abril, 2008)
Resumen
En este estudio, se analizó la relación de la variación de peso corporal (PC) y de condición corporal (CC), con el tiempo de aparición de los primeros folículos dominantes (FD) y, la presencia de receptores de leptina y de LH en los ovarios. En once vacas cebú se realizó ultrasonografía ovárica durante el posparto para determinar su dinámica folicular: tan pronto como el tercer folículo dominante (FD) fue detectado, las vacas fueron tratadas con LH (FDLH, n = 4) o solución salina (grupo control, FDSS, n = 4) y, 48h después, fueron sometidas a ovariectomía para determinar receptores de leptina y de LH por inmunofluorescencia indirecta (IFI) e immunohistoquímica (IHQ). Las vacas del grupo de control que no presentaron FD también fueron ovariectomizadas (n = 3). No se encontró ningún efecto significativo del PC y la CC sobre los días al tercer FD. Sin embargo, se halló una correlación negativa entre la pérdida de PC y la pérdida diaria de CC y el tiempo al primer y tercer FD. En el grupo FDLH, el 75% (3/4) de las vacas tratadas, ovularon, mientras que en el grupo FDSS, ninguna ovuló. La IFI no fue técnicamente posible, ya que se encontró autofluorescencia en los tejidos. En la IHQ para ambos receptores, se encontró un patrón de expresión similar durante el desarrollo folicular (lo que sugirere una comodulación en el ovario), cuya expresión fue mayor en los FD y en cuerpos hemorrágicos y menor en folículos en crecimiento, tanto en las células de la granulosa, como de la teca y en células luteinizadas.
Palabras clave: anestro posparto, receptor de leptina, receptor de LH
Summary
This study seek for the relationship between body weight (BW) and body condition, the time of appearance of first dominant follicles (DF) and the presence of leptin and LH receptors in the ovaries. Ovarian ultrasonography was performed during the postpartum period on eleven Zebu cows to monitor their follicular dynamics. When the third DF was detected, the cows were treated with LH (n=4) or saline solution (n=4) and 48h later they were ovariectomized to determine leptin and LH receptors by indirect immunofluorescence (IFI) and immunohistochemistry (IHQ). Cows that did not display DF were also ovariectomized (control, n=3). No significant effect of variation of BW and BC on the days to third DF was found. There was also a negative correlation between BW loss and daily BW loss with the time to the first and third DF. In FDLH group, 75% (3/4) of cows, ovulation was induced. In FDSS group, cows did not ovulate. The IFI studies for leptin and LH receptors detection were technically impossible due to the presence of autofluorescence. In the IHQ studies for both receptors, pattern of expression similar during the follicular development was found suggesting a comudulation at the ovarian level, being greater in hemorrhagic bodies and dominant follicles, and minor in growing follicles, at granulosa, theca and luteinized cells levels.
Key words: leptin receptor, LH receptor, postpartum anestrus
]]>Resumo
Neste estudo, analisaram a relação entre a variação no peso corporal (PC) ea BCS (CC), com o tempo de aparecimento dos primeiros folículos dominante (FD), e na presença de leptina e receptores de LH ovários. Em onze vacas zebus foi realizado durante o pós parto ultra-som para determinar a sua dinâmica folicular: logo que o terceiro folículo dominante (FD) foi detectado, as vacas foram tratadas com LH (FDLH, n = 4) ou soro fisiológico (grupo controle, FDSS, n = 4) e 48 horas mais tarde, veio a determinar nos termos ovariectomía receptores de leptina e LH por imunofluorescência indireta (IFI) e immunohistoquímica (IHQ). Vacas no grupo de controlo que não apresentarem FD também foram ovariectomizadas (n = 3). Nós não encontramos qualquer efeito significativo sobre o PC do CC no dia de terceiro FD. No entanto, encontramos uma correlação negativa entre a perda do PC e da perda diária de CC e da hora em primeira e terceira FD. Do grupo FDLH-tratados, 75% (3/4) vacas ovularam. Do grupo FDSS, nenhum ovo. O IF não era tecnicamente possível, uma vez que autofluorescencia foi encontrada nos tecidos. No IHQ para ambos os receptores, foi encontrado um padrão semelhante de expressão durante o desenvolvimento folicular sugerindo uma comodulación no ovário, cuja expressão foi maior no DF e corpo hemorrágico nas instâncias inferiores e de folículos em crescimento, tanto nas células granulosa, como nas células da teca e células luteinizadas.
Palavras chave: anestro pós parto, receptor leptina, receptor LH
Introducción
El ganado Cebú (Bos indicus indicus) destinado a la producción de carne en el trópico bajo colombiano, con amamantamiento permanente, presenta una baja eficiencia reproductiva, debido principalmente a su prolongado anestro posparto (8 a 10 meses posparto) (18). El papel de la hormona luteinizante (LH) en el anestro posparto bovino ha sido reportado en la literatura desde hace más de tres décadas (9, 10). La leptina (OB) fue descubierta a mediados de los noventas (40) y su importancia en la fisiología reproductiva ha sido ampliamente reportada (2). Sin embargo, los mecanismos celulares y moleculares en las gónadas y las vías de regulación y señalización implicados en la regulación del anestro posparto bovino, todavía no están completamente entendidos.
El descenso rápido en las concentraciones séricas de estradiol al parto, retira el bloqueo ejercido por esta hormona sobre el eje hipotálamo-hipófisis durante la gestación, estimula la síntesis de ARNm para la producción de gonadotropinas y aumenta la liberación de hormona liberadora de gonadotropinas (GnRH) (26). Las células gonadotrópicas de la adenohipófisis llenan sus gránulos de LH, pero, aunque dicha hormona es necesaria para la maduración folicular y la ovulación, no alcanza las concentraciones séricas necesarias para inducir la ovulación, debido al efecto del vínculo entre la vaca y el ternero (16, 33, 36), a la frecuencia de succión del ternero (28, 35, 36) y a los cambios en condición corporal (4, 18), todos los cuales inducen una liberación pulsátil de baja frecuencia y baja amplitud.
]]> Durante el ayuno, por su parte, la señal de la OB es removida, y la pulsatibilidad de la LH y la función reproductiva declinan rápidamente; de ahí que el ayuno causa cesación del ciclo estral en ratones (39) y en bovinos (4), el cual puede ser restaurado por la realimentación o administración de la OB durante dos días, evento que es acompañado por un incremento de la LH plasmática.Para realizar la esteroidogénesis, la expresión de los receptores para el ingreso de colesterol y el transporte del colesterol intracitoplasmático, son controlados, entre otros, por un factor de transcripción conocido como la proteína reguladora de esterol (SREBP, que posee un elemento de unión a los elementos SRE de los promotores de algunos genes implicados en la esteroidogénesis), sobre el cual la OB tiene actividad reguladora (27). Luego, este colesterol citoplasmático sirve como sustrato para la producción consecutiva de pregnenolona, progesterona, 17α-hidroxiprogesterona y androstenediona, a través de una cascada enzimática regulada por la interacción de la LH con su receptor en las células de la teca; la producción de estrógenos se incrementa a medida que se da el crecimiento de las ondas foliculares y, en la luteinizacion, se cambia la ruta metabólica hacia la producción de progesterona (12, 15, 20). La identificación de las sustancias inhibitorias responsables de la disminución en la concentración de FSH y de las sustancias involucradas en el cambio a la dependencia de LH durante la divergencia y la dominancia folicular no han sido bien clarificadas (13).
Las vías de señalización en la dependencia a LH en las células de la teca producen cambios en la expresión de genes que son críticos para la producción de estrógenos; específicamente, la LH estimula la producción de altos niveles de androstenediona. Además, el estado nutricional del animal es un factor importante en la regulación de la esteroidogéneis, donde la OB tiene un papel clave, esta, producida en el tejido adiposo, participa en vías de señalización para regular el balance energético, por parte del hipotálamo, expresado en la cantidad de grasa corporal. La expresión de diferentes isoformas de OB-R se ha demostrado en el humano, ratones, ratas, bovinos, caprinos, suinos, y en gran variedad de tejidos como los riñones, el hígado, el bazo, el cerebro, y el aparato reproductivo (5, 27).
Otras hormonas como la insulina y el IGF-I también regulan la función reproductiva en términos del estado energético del animal. La insulina tiene un efecto estimulador de la esteroidogénesis, al inducir la actividad de la aromatasa en la granulosa, efecto que puede ser antagonizado directamente por la OB especialmente en las células indiferenciadas, mostrando que el número de receptores para OB en los folículos maduros es menor y por lo tanto es menos sensibles al efecto negativo de la OB; además, en ausencia de insulina, la OB tiene poco o ningún efecto en la esteroidogénesis (31). Sin embargo, el efecto en las células de la granulosa de bovinos no parece ser una inhibición directa de la unión de la insulina a su receptor y no afecta el crecimiento basal de éstas células mediado por insulina (31), aunque sí lo hace disminuyendo el crecimiento mediado por IGF-I (32). Más aún, la OB no tiene efectos en la inducción de la producción de estradiol mediada por IGF-I en las células de la granulosa de bovinos, en la producción de progesterona mediada por FSH en las células de la granulosa de ratas, ni en la producción de estradiol y progesterona mediada por LH en ovarios de ratas. Por su parte, varios autores sugieren que las altas concentraciones de OB en mujeres obesas inhiben la esteroidogénesis de la granulosa mediada por IGF-I, insulina o gonadotropinas (11, 32).
Nuevos estudios son necesarios para aclarar la acción inhibitoria de la OB en el ovario bovino, de rata y humano. En las células de la teca en bovinos, la OB inhibe la esteroidogénesis mediada por insulina —aunque, igual que en la granulosa, no parece ser de forma directa y la producción de androstenediona mediada por LH en humanos (32). Por consiguiente, en este estudio se evaluó la capacidad ovulatoria de los primeros folículos dominantes y el efecto de los cambios de peso y condición corporal en la dinámica folicular posparto y su relación con la expresión de receptores de OB y de LH en los folículos ovulados y no ovulados.
Materiales y métodos
Localización del estudio
El estudio fue realizado en la Hacienda “El Progreso”, propiedad de la Universidad de Antioquia, ubicada en el municipio de Barbosa (El Hatillo, Antioquia, Colombia), localizada en una formación ecológica de bosque húmedo premontano (bh-PM) según la clasificación de Holdridge (19), a 1400 msnm, con una temperatura promedio de 20 °C, y una precipitación anual de 1300 mm. Los animales pastaron en potreros sembrados en pasto nativo, pasto estrella (Cynnodon spp) pasto uribe (Hypharrenia ruffa) y pasto braquiaria (Brachiaria humidicola), con disponibilidad de agua corriente y sal mineralizada (con un contenido de fósforo del 8%) a voluntad.
Unidades experimentales
]]> En la región, fueron seleccionadas 11 vacas cebú en posparto, de tres a siete lactancias, una condición corporal igual o superior a 3.0 (Escala de 1 a 5) y en estado de preñez avanzada. Los animales pastorearon a voluntad y al momento del parto no se apartaron de sus terneros.Grupos y tratamientos
Una semana después del parto, se dio inicio al seguimiento de la dinámica folicular cada 48 horas mediante ecografía transrectal (Aloka SSD-500, tiempo real, sonda 5MHz, Aloka CO, LTD, Tokio, Japón). Los diámetros foliculares y la ubicación de los folículos fueron registrados, y se determinó como folículo dominante aquel que presentaba un diámetro superior a 10 mm durante dos observaciones consecutivas. Los grupos se conformaron de la siguiente manera: 1) grupo folículo dominante–tratamiento con solución salina (FDSS, n = 4), donde las vacas fueron tratadas con 2 ml de solución salina al 5%, vía intravenosa, al observarse el tercer folículo dominante; 2) grupo folículo dominante–tratamiento con hormona luteinizante (FDLH, n = 4), donde las vacas fueron tratadas con 3.000 UI de hCG (Chorulon® , Intervet, Netherlands) vía intravenosa, al observarse el tercer folículo dominante. En ambos grupos se realizó ecografía ovárica a las 24 y 48 horas para determinar si ovulaban o no y a las 48 horas del tratamiento se realizó ovariectomía; y 3) grupo control (CON, n = 3), donde las vacas que no presentaron dominancia folicular antes de 60 días, no recibieron ningún tratamiento y se les realizó ovariectomía. El peso de los animales se midió utilizando cinta bovinométrica (17), y la condición corporal fue evaluada en escala de 1 a 5 (28), ambos, una vez por semana.
Procedimiento de ovariectomía
Ambos ovarios de cada animal fueron extraídos por ovariectomía lateral izquierda. Las vacas fueron sedadas y se les aplicó anestesia paravertebral para el procedimiento quirúrgico. Luego, los ovarios fueron transportados en solución salina fosfatada (PBS) a 4 °C, y luego fueron congelados y conservados a -20 °C hasta su procesamiento en el laboratorio.
Detección de receptores por inmunofluorescencia e inmunohistoquímica
Las estructuras de los ovarios fueron clasificadas como folículos en crecimiento, folículo dominante y cuerpo hemorrágico. Los cortes se embebieron en Optimal Cutting Temperature (OCT compound® , Tissue-Tek, Sakura USA, Torrance, CA) y se realizaron criocortes de 5µ, que se montaron en láminas de vidrio tratadas con polilisina. Los tejidos fueron congelados a -20°C hasta que fueron procesados. Las muestras se fijaron en acetona fría (4ºC/10 min), luego se realizaron lavados consecutivos con TBS y se recuperó el antígeno colocándolas en un tampón de citrato (0.01 M, pH 6.0) y sometiéndolas a dos ciclos de microondas de 15 minutos (máxima temperatura) cada uno. El bloqueo de sitios inespecíficos se hizo con albúmina sérica bovina al 5% en PBS a temperatura ambiente/60 min. Como primer anticuerpo se utilizaron: anticuerpo policlonal de ratón dirigido contra el receptor de leptina humano (con 86% de homología con la proteína bovina; Sigma, USA, Product Nº L9411) diluido 1:20 e incubado a 37 ºC/2 horas; o anticuerpo policlonal de cabra dirigido contra el receptor de LH caprino, el cual presenta un 96% de homología con la proteína bovina (Santa Cruz Biotechnology, Inc. USA; LHR K-15: sc-26341), diluido 1:20 a 37 ºC/1 hora. Para la inmunofluorescencia se utilizaron como segundos anticuerpos: anticuerpo monoclonal de cabra dirigido contra los fragmentos pesado y ligero de la región constante de IgG de ratón (anti-anti-receptor de leptina) conjugado con CY3 (Sigma-Aldrich, Inc. USA, C2821), diluido 1:200 a 37 ºC/2 horas; y anticuerpo monoclonal de cabra dirigido contra los fragmentos pesado y ligero de la región constante de IgG de ratón (anti-anti-receptor de LH), conjugado con FITC (Sigma-Aldrich, Inc. USA, F2016), diluido 1:200, a 37 ºC/2 horas. Además, se realizó una tinción de contraste con DAPI (5 µg/ml). Para la inmunohistoquímica se utilizaron como segundos anticuerpos: anticuerpo monoclonal de conejo dirigido contra Ig de ratón y cabra (anti-anti-receptor de leptina o de LH, Invitrogen®, R21455, R21459), conjugados con peroxidasa. La actividad de la peroxidasa endógena de los tejidos se bloqueó con peróxido de hidrógeno (al 3% en metanol)/5 min, y luego se reveló la actividad de la peróxidasa de rábano con diaminobencidina (0.5 mg/ml; en TBS; DAB, Sigma). Finalmente, se realizó una tinción de contraste con hematoxilina.
Como control positivo de OBR se utilizaron cortes de hipotálamo bovino, y como control positivo de LHR células de granulosa bovina cultivadas in vitro o cortes de folículo dominante de una vaca ciclando (control positivo); como control negativo se usaron cortes de epidermis bovina (control negativo OBR y LHR). Además, se realizaron los controles negativos de la técnica, sin agregar el primer anticuerpo durante el procesamiento.
Luego del procesamiento de las muestras, los cortes fueron analizados de la siguiente manera: 1) de los campos representativos para cada estructura ovárica se tomaron fotografías a 40X; 2) las fotografías fueron descargadas en el programa Adobe Photoshop® (Ver. 4.0, Adobe Systems); 3) una máscara de dimensiones fijas se seleccionó para todas las muestras, la que se desplazaba en las zonas de interés (células de la granulosa, teca y luteinizadas), para contar en su interior, el número de células positivas (con expresión de cada uno de los receptores) y el número de células negativas (sin expresión), hasta ajustar un total de 100 células/campo; y 4) los datos fueron tabulados, se hallaron los porcentajes de expresión de cada receptor en cada estructura y, se procesaron para estadística descriptiva, para obtener los promedios y las desviaciones estándar.
Medición de estrógenos séricos por radioinmunoanálisis
]]> Muestras de suero de cada uno de los animales se tomaron una vez a la semana y en el momento de la ovariectomía; el suero fue procesado por radioinmunoensayo (coeficiente de variación intraensayo de 4.85%) para determinar las concentraciones de 17β-estradiol en cada una de las muestras.Análisis estadístico
Para estudiar la capacidad ovulatoria del tercer folículo dominante, se analizaron los datos de forma descriptiva, agrupando las vacas que presentaron dominancia folicular (FDSS, FDLH, n = 8). En este mismo grupo, se realizó una correlación de Pearson entre la variación en la condición corporal y el peso corporal, con los días a la dominancia folicular. Para el análisis de la expresión de OBR y LHR, se tomaron los promedios de expresión en cada una de las estructuras (folículo en crecimiento, folículo dominante y cuerpo hemorrágico) y se les realizó un Anova para un diseño desbalanceado. La homogeneidad de varianzas se evaluó mediante prueba de Levene. Además, se realizaron correlaciones de Spearman entre la expresión de receptores y las variables de cambio en la condición corporal, el peso y los días a la dominancia folicular. Todos los análisis estadístico se realizaron utilizando el programa Statistica® (Ver. 5.0, StatSoft, USA). Para todos los efectos se estimó una diferencia significativa de p<0.05.
Resultados
Reanudación de la dinámica folicular posparto y variación en la CC y el peso
En el posparto temprano, las vacas presentaron un peso de 366.42 ± 41.89 kg y una CC de 3.5 ± 0.25 (promedio ± desviación estándar). Hasta los 30 días posparto el peso disminuyó en un índice de 0.68 ± 1.32 kg/día, con vacas que perdieron desde 1g hasta 50 kg en el período de seguimiento folicular; mientras que, la CC se redujo hasta 2.5 ± 0.45. Luego de una semana del parto todas las vacas ingresaron a seguimiento de la dinámica folicular y, al momento de la primera evaluación todas presentaron cohortes de folículos en fase de emergencia. En el grupo de vacas que presentaron dominancia folicular (FDLH y FDSS, n=8), la primera y tercera dominancia folicular ocurrieron a los 29.5 ± 14.6 y 53.6 ± 17 días, respectivamente. Las vacas control (n = 3) fueron ovariectomizadas a los 35 ± 6.7 días posparto, sin haber presentado FD.
Capacidad ovulatoria del tercer folículo dominante posparto
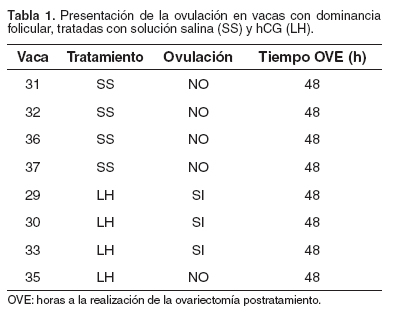
En el grupo FDLH, de las cuatro vacas tratadas, en tres (75%) se indujo la ovulación del tercer folículo dominante, luego de la aplicación exógena de LH. En el grupo FDSS, ninguna de las vacas ovuló, luego de la aplicación de solución salina (véase Tabla 1).
Correlación entre la variación de la condición corporal, el peso corporal y los días a la dominancia folicular
]]> De forma general, incluyendo todos los animales que presentaron dominancia folicular, no se encontró un efecto significativo (p>0.05) entre la variación de la CC y PC, sobre la presentación de la dominancia folicular, incluso cuando en el análisis fueron incluidas las vacas con marcada pérdida de PC. De igual modo, en los análisis de regresión se encontró una asociación negativa no significativa (r = -0.45, R2 = 20.25%, p>0.05) entre la pérdida de peso diaria y la presentación de dominancia y, una asociación positiva no significativa (r = 0.25, R2 = 6.35%; p>0.05) entre el cambio de la condición corporal y los días a la presentación de la primera dominancia folicular. Estas mismas asociaciones, entre pérdida de peso diaria (r = -0.38, R2 = 14.44%; p>0.05), pérdida de condición corporal (r = 0.19, R2 = 3.61%; p>0.05) se hallaron al día de aparición de la tercera dominancia folicular. Al tomar este mismo grupo con dominancia folicular y hacer una correlación entre la pérdida de peso diaria y los días a la primera y tercera dominancia folicular, se encontró que la pérdida de peso diaria determinaría en un 26% los días a la aparición del primer folículo dominante (r = -0.51) y, en un 37% los días al tercer folículo dominante (r = -0.61) (p<0.05).
Determinación de la expresión de receptores de leptina y de LH por inmunofluorescencia
La determinación de la expresión de OBR y LHR por IFI, no mostró resultados concluyentes debido a que el tejido ovárico siempre presentó autofluorescencia, lo que impedía encontrar un patrón de expresión. Para contrarrestar este fenómeno se realizaron varios tratamientos: 1) aumento de lavados con PBS, Tritón-X y Tween-20; 2) bloqueo con BSA al 10%, leche descremada al 5%, o suero canino (Dr. Carlos Vélez y Dra. Marlene Jiménez, Universidad de Antioquia, comunicación personal), y 3) exposición a lámpara de mercurio/6 h y a lámparas UV (20W) y Neón (18W)/48 h (24). En ninguno de los casos se obtuvo éxito.
Determinación de la expresión de receptores de leptina y de LH por inmunohistoquímica
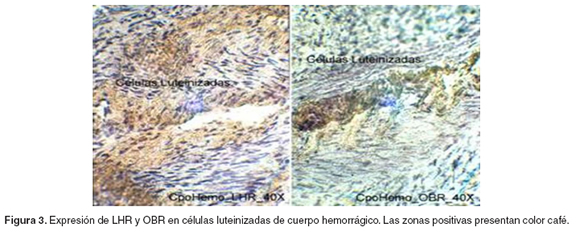
La IHQ reveló la presencia de OBR y LHR en las células luteinizadas de los cuerpos hemorrágicos y en células de la granulosa y de la teca (véanse Figuras 1-3).
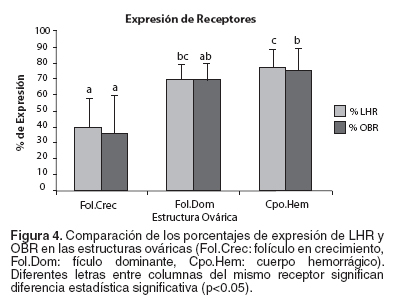
Con respecto a la expresión de LHR, se encontró un patrón de expresión bajo en los folículos en crecimiento (p<0.05), comparado con la alta expresión en los folículos dominantes y en los cuerpos hemorrágicos, entre los que no hubo diferencia significativa. La expresión de OBR presentó la misma tendencia que LHR, aunque sólo hubo diferencia significativa entre la expresión en folículos en crecimiento y en los cuerpos hemorrágicos (p<0.05) (véase Figura 4).
]]>
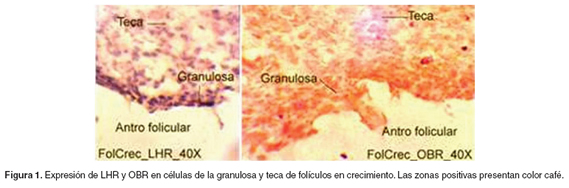
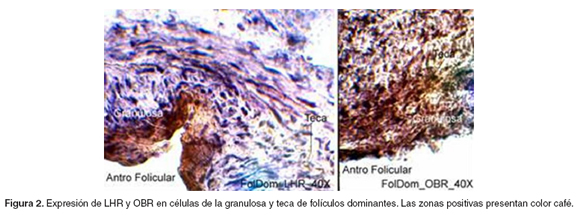
Determinación de los niveles séricos de 17β-estradiol
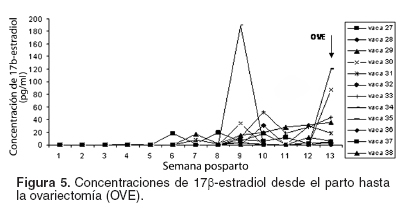
Todas las vacas presentaron niveles séricos basales de 17β-estradiol desde el parto hasta la presentación de la primera dominancia folicular (véase Figura 5). De la misma manera, todas las vacas de los grupos FDSS y FDLH presentaron niveles altos de estrógenos circulantes. Sin embargo, no se encontró una correlación significativa entre estos resultados y los días a la dominancia folicular, la presentación de receptores de ambas hormonas o la capacidad ovulatoria de los animales.
]]>
Discusión
Durante el posparto temprano, las vacas cebú presentan actividad folicular temprana, ya que se observan folículos emergentes relacionados con el crecimiento de cohortes foliculares, alrededor de una semana luego del parto. En este estudio, incluso, se pudo observar que la mayoría de las vacas (FDLH y FDSS, n = 8) presentaron dominancia folicular temprana (alrededor de un mes posparto), con folículos que tienen capacidad ovulatoria ya que responden a la administración de análogos de LH. Por lo tanto, el fenómeno de la anovulación posparto puede ser debido a la incapacidad para la producción de un pico preovulatorio de LH, más que una disminución en su frecuencia de liberación: la vaca hace dominancia folicular pero sólo ovula si tiene un aumento rápido y significativo de la LH circulante. La ausencia del pico preovulatorio de LH parece deberse al efecto del amamantamiento permanente que tenían los animales experimentales, de acuerdo con lo reportado en la literatura (14, 18), donde la repleción de la adenohipófisis con LH ocurre en las primeras semanas posparto en las vacas, a pesar de que no ocurre la liberación del pulso de LH necesario para la maduración folicular y ovulación, debido al efecto del amamantamiento del ternero (36). Luego del parto, las vacas cebú reanudan pronto su actividad ovárica, entre 26 a 78 días posparto, pero el anestro y la anovulación se extienden hasta los 7 a 12 meses posparto, tiempo que generalmente coincide con el destete de la cría (28). Ésta condición es definida por Wiltbank et al (37) como “anovulación con crecimiento folicular únicamente hasta la emergencia”. De conformidad con los hallazgos del presente estudio, este fenómeno se ve relacionado, al parecer, con la expresión de LHR en las células de la granulosa y de la teca, lo que apoya el hecho de que las vacas sí tienen la capacidad ovulatoria —entendiendo que presentan el primer suceso necesario para la ruta de señalización para la ovulación, pero que no se desencadena por ausencia de ligando: la LH en forma de pico preovulatorio.
Ni la pérdida del peso ni la variación en la CC fueron factores determinantes en la presentación de la primera dominancia folicular, lo que concuerda con otros estudios donde el peso corporal no fue el factor limitante para la presentación del estro (22), al igual que con otras investigaciones en las que la CC posparto no tuvo tanta importancia en la fisiología reproductiva, como lo tuvieron la CC y el peso preparto (8, 25, 38). Por el contrario, a diferencia de otros estudios donde la variación en CC y PC fueron variables que mostraron una influencia clara en la reanudación de la dinámica folicular y en la establecimiento de la dominancia folicular posparto, en nuestro estudio no presentaron un impacto significativo. Por otra parte, se observaron correlaciones entre el período a la presentación de las primeras dominancias foliculares más corto y la mayor pérdida de peso de la vaca. Sin embargo, esto se tradujo en que el momento de presentación de los primeros folículos dominantes explica sólo en un 20.25% las variaciones de PC y de CC, lo que pone en evidencia que el fenómeno es de tipo multifactorial. Además, las vacas no presentaron ovulación espontánea durante el periodo de estudio, lo que permite sugerir que en el microambiente folicular se encuentren las condiciones de señalización para OB y LH por la expresión de sus receptores, pero, las concentraciones circulantes de sus respectivos ligandos, que varían según el estado nutricional de la vaca, son un factor clave para inducir la ovulación.
La expresión de LHR fue significativamente mayor en las vacas que presentaron dominancia folicular, hubieran ovulado o no, comparado con las vacas que no hicieron dominancia folicular y sólo presentaban folículos en crecimiento. Durante el período de dominancia folicular se producen altas cantidad de estrógenos (especialmente 17β-estradiol), los cuales son necesarios para la ovulación. El pico preovulatorio del pulso de LH ocurre luego del pico de estradiol (13), por lo tanto, la ausencia de este en la anovulación posparto, puede deberse a una disminución en la esteroidogénesis la cual está regulada tanto por LH como por OB.
En este estudio se reporta, por primera vez, la expresión de receptores para OB en las células de la teca, la granulosa, y en células luteinizadas de ovario bovino. En ovario de ratas, se reportó la expresión del OBR en los oocitos, células endoteliales, de la teca y el cuerpo lúteo (7, 29), donde el porcentaje de expresión fue menor en los folículos en crecimiento y aumentó proporcionalmente con el desarrollo folicular. En gallinas también se encontró un patrón de expresión del OBR (3), que fue constante en células de la teca, pero disminuyó en las células de la granulosa con la diferenciación folicular, en gallinas de crecimiento lento. En murinos también se halló una alta expresión en células de la teca, y menor, en oocitos, células de la granulosa y células luteinizadas (30). En porcinos se halló un patrón de expresión en células de la teca, granulosa y células luteinizadas, el cual aumentó en paralelo con el progreso de la luteinizacion (27). En humanos, se encontró expresión del OBR en los oocitos y las células estromales (1, 6, 21) y en el líquido folicular (34). De forma general, estos hallazgos sugieren el papel que tiene la señalización por leptina en el ovario, para los procesos de esteroidogenesis y maduración de oocitos.
En el presente cambio, los OBR se observaron con una expresión similar a los LHR, durante los estadios consecutivos de crecimiento folicular y luteinizacion temprana, en cada una de las estructuras ováricas evaluadas. Este hallazgo sugiere un papel importante de la relación OB-LH y su comodulación positiva sobre la expresión de receptores foliculares, durante la esteroidogenesis en bovinos, y la adquisición de la capacidad ovulatoria para resolver el anestro posparto. Por otro lado, en un modelo de luteinización in vitro desarrollado para hallar la expresión de ambos receptores (23), se encontró que la comodulación entre ambos receptores es negativa; esto es, que la expresión máxima de un receptor coincide con la expresión mínima del otro y que además esta interacción parece depender del grado de luteinización de las células de la granulosa en cultivo, lo que sugiere que estos dos modelos se complementan y permiten describir la dinámica de ambos receptores desde el desarrollo folicular, la ovulación y la formación del cuerpo lúteo. Poco antes de la ovulación, las células de la granulosa y la teca comienzan un proceso de luteinización aumentando la síntesis y liberación de progesterona, la cuál alcanza niveles altos en suero luego de la ovulación y formación del cuerpo lúteo, por parte de las células de la granulosa y de la teca agrupadas.
Aparentemente la disminución en la secreción de OB, y en su ARNm, en la fase folicular temprana y tardía, no guarda relación con la producción de progesterona, en contraste con resultados obtenidos en mujeres y roedores en los cuales la OB sérica aumentó luego de una terapia de reemplazo con estradiol y progesterona. En cultivos de células de granulosa y la teca de cerdos y en cortes de ovario en diferentes estadios de la fase lútea, OB y su receptor tuvieron alta concentración en el CL de mitad de ciclo y baja en el CL posovulatorio y en regresión, lo que indicaría una regulación positiva de la OB en la producción de progesterona (27).
]]> El estudio permite concluir que en el fenómeno fisiológico del anestro posparto bovino, las vacas presentan una reactivación ovárica temprana y llegan a adquirir rápidamente su capacidad ovulatoria, entendida por la expresión de receptores para la LH y la leptina en los folículos, pero esta se ve limitada por la pérdida de peso diaria y por la ausencia del pico preovulatorio de LH. Ambas hormonas, se comodulan en el ovario y regulan la esteroidogénesis, ya que se pudo observar un patrón de expresión similar de receptores que aumentaba a medida que avanzaba el desarrollo folicular. Estos hallazgos permiten dar el primer paso en el modelo hipotético de esteroidogénesis planteado, pero, es necesario realizar investigaciones adicionales encaminadas a completar la ruta de señalización de los receptores para ambas hormonas, para establecer a cuales genes regulan su expresión en el ovario y correlacionar su expresión con niveles séricos para las hormonas esteroideas.
Agradecimientos
Este proyecto fue financiado por la Internacional Foundation for Science (IFS). Al Departamento de Formación Académica de Haciendas, de la Facultad de Ciencias Agrarias de la Universidad de Antioquia por el préstamo de las instalaciones de la Hacienda El Progreso. A la Dra. Doris Ruíz del Departamento de Dermatología del Hospital San Vicente de Paúl por su colaboración en el procesamiento de los cortes de tejido. Al Dr. Henry Grajales del Laboratorio de Radioinmunoanálsis en el Departamento de Producción Animal de la Universidad Nacional de Colombia (Sede Medellín), por su colaboración en el procesamiento de los sueros para la medición de estrógenos.
Referencias
1. Abir R, Ao A, Jin Sh, Barnett M, Raanani H, et al. Leptin and its receptors in human fetal and adult ovaries. Fertil Steril 2005; 84:1779-1782. (Abstract) [ Links ]
2. Barash IA, Cheung CC, Weigle DS, Ren H, Kabigting EB, et al. Leptin is a metabolic signal to the reproductive system. Endocrinology 1996; 137:3144-3147. (Abstract) (Pdf) [ Links ]
3. Cassy S, Metayer S, Crochet S, Rideau N, Collin A, et al. Leptin receptor in the chicken ovary: potential involvement in ovarian dysfunction of ad libitum-fed broiler breeder hens. Reprod Biol Endocrinol 2004; 2:72-77. (Html) (Pdf) [ Links ]
4. Castillo GH, Ruíz ZT, Olivera M, Jiménez C. Reactivación ovárica posparto en vacas Cebú Brahman con relación al peso y la condición corporal. Rev Colomb Cienc Pecu 1997; 10:12-18. [ Links ]
5. Chilliard Y, Bonnet M, Delavaud C, Faulconnier Y, Leroux C, et al. Leptin in rumiants. Gene expression in adipose tissue and mammary gland, and regulation of plasma concentration. Domest Anim Endocrinol 2001; 21:271-295. (Abstract) (Html) [ Links ]
6. Cioffi JA, Van Blerkom J, Antczak M, Shafer A, Wittmer S, et al. The expression of leptin and its receptors in pre-ovulatory human follicles. Mol Hum Reprod 1997; 3:467-472. (Abstract) (Pdf) [ Links ]
7. Duggal PS, Weitsman SR, Magoffin DA, Norman RJ. Expression of the long (OB-RB) and short (OB-RA) forms of the leptin receptor throughout the oestrous cycle in the mature rat ovary. Reproduction 2002; 123:899-905. (Abstract) (Pdf) [ Links ]
8. Dunn TG y Kaltenbach CC. Nutrition and the postpartum interval of the ewe sow and cow. J Anim Sci 1980; 51 Supl 2:29. (Abstract) [ Links ]
9. Echternkamp SE, Hansel W. Concurrent changes in bovine plasma hormone levels prior to and during the first postpartum estrous cycle. J Anim Sci 1973; 37:1362-1370. (Abstract) (Pdf) [ Links ]
10. Edgerton LA, Hafs HD. Serum luteinizing hormone, prolactin, glucocorticoid, and progestin in dairy cows from calving to gestation. J Dairy Sci 1973; 56:451-458. (Abstract) (Pdf) [ Links ]
11. Erickson GF, Shimasaki S. The physiology of folliculogenesis: the role of novel growth factors. Fert Steril 2001; 76-82. (Abstract) [ Links ]
12. Evans ACO, Fortune JE. Selection of the dominant follicle in cattle occurs in the absence of differences in the expression of messenger ribonucleic acid for gonadotropin receptors. Endocrinology 1997; 138:2963-2971. (Abstract) (Html) (Pdf) [ Links ]
13. Ginther OJ, Wiltbank MC, Fricke PM, Gibbons JR, Kot K. Selection of the dominant follicle in cattle. Biol Reprod 1996; 55:1187-1194. (Abstract) (Pdf) [ Links ]
14. Giraldo CA, Ruíz ZT, Restrepo LF, Olivera M. Interrupción temporal del amamantamiento (ITA) en vacas Cebú y su efecto en la función ovárica. Redvet 2005; VI Diciembre. http://www.veterinaria.org/revistas/redvet/n121205/120502.pdf [ Links ]
15. Gore-Langton RE, Armstrong DT. Follicular steroidogenesis and its control. In: Knobil E, Neill JD (eds). The Physiology of reproduction. Vol 1, 2a ed. New York: Raven Press; 1998. p571-627. [ Links ]
16. Griffith MK, Williams GL. Roles of maternal vision and olfaction in suckling-mediated inhibition of luteinizing hormone secretion, expression of maternal selectivity, and lactational performance of beef cows. Biol Reprod 1996; 54:761-768. (Abstract) (Pdf) [ Links ]
17. Heinrichs J, Lammers B. 2001. Monitoring dairy heifer growth. Pennsylvania State Univer. http://www.das.psu.edu/dcn/calfmgt/growth/docs/measure.html [ Links ]
18. Henao G, Olivera-Ángel M, Maldonado-Estrada JG. Follicular dynamics during the postpartum anestrous and the first estrous cycle in suckled or non-suckled Brahman (Bos indicus) cows. Anim Reprod Sci 2000; 63:127-136. (Abstract) [ Links ]
19. Holdriege LR. Ecología basada en zonas de vida. Instituto interamericano de cooperación para la agricultura. San José de Costa Rica. 1996. [ Links ]
20. Ireland JJ, Mihm M, Austin E, Diskin MG, Roche F. Historical perspective of turnover of dominant follicles during the bovine estrous cycle: key, concepts, studies, advancements, and terms. J Dairy Sci 2000; 83:1648-1658. (Abstract) (Pdf) [ Links ]
21. Karlsson C, Lindell K, Svensson E, Bergh C, Lind P, et al. Expression of functional leptin receptors in the human ovary. J Clin Endocrinol Metab 1997; 82:4144-4148. (Abstract) (Html) (Pdf) [ Links ]
22. Mejía C, Henao G, Botero J, Acevedo LI, Giraldo AM, et al. Variaciones en el peso y la condición corporal posparto y su relación con algunos parámetros de eficiencia reproductiva en vacas cebú. Rev Colomb Cienc Pecu 2003; 16 Supl: 85. (Abstract) (Html) (Pdf) [ Links ]
23. Montaño EL, Ruíz-Cortés ZT. Expresión de receptores de leptina y de LH; efecto de sus ligandos en la esteroidogénesis en un modelo de luteinización in vitro. Maestría en Ciencias Animales, Universidad de Antioquia, Trabajo de grado. 2006. [ Links ]
24. Neumann M, Gabel D. Simple method for reduction of autofluorescence in fluorescence microscopy. J Histochem Cytochem 2002; 50:437-439. (Abstract) (Html) (Pdf) [ Links ]
25. Richards MW, Spitzer JC, Warner MB. Effect of varying levels of postpartum nutrition and body condition at calving on subsequent reproductive performance in beef cattle. J Anim Sci 1986; 62:300-305. [ Links ]
26. Robinson JJ. Nutrition and reproduction. Anim Reprod Sci 1996; 42:25-34. [ Links ]
27. Ruíz-Cortés ZT, Martel-Kennes Y, Gevry NY, Downey BR, Palin MF, et al. Biphasic effects of leptin in porcine granulosa cells. Biol Reprod 2003; 68:789-796. (Abstract) (Html) (Pdf) [ Links ]
28. Ruíz-Cortés ZT, Olivera-Ángel M. Ovarian follicular dynamics in sucked zebu (Bos indicus) cows monitored by real time ultrasonography. Anim Reprod Sci 1999; 54:211-220. (Abstract) [ Links ]
29. Ryan NK, van der Hoek KH, Robertson SA, Norman RJ. Leptin and leptin receptor expression in the rat ovary. Endocrinology 2003; 144:5006-5013. (Abstract) (Html) (Pdf) [ Links ]
30. Ryan NK, Woodhouse CM, van der Hoek KH, Gilchrist RB, Armstrong DT, et al. Expression of leptin and its receptor in the murine ovary: possible role in the regulation of oocyte maturation. Biol Reprod 2002; 66:1548-1554. (Abstract) (Html) (Pdf) [ Links ]
31. Spicer LJ, Francisco CC. The adipose obese gene product, leptin: evidence of a direct inhibitory role in ovarian function. Endocrinology 1997; 138:3374-3379. (Abstract) (Html) (Pdf) [ Links ]
32. Spicer LJ. Leptin: a possible metabolic signal affecting reproduction. Domestic Anim Endocrinol 1991; 21:257-270. (Abstract) [ Links ]
33. Stagg K, Spicer LJ, Sreenan JM, Roche JF, Diskin MG. Effect of calf isolation on follicular wave dynamics, gonadotropin and metabolic hormone changes, and interval to first ovulation in beef cows fed either of two energy levels postpartum. Biol Reprod 1998; 59:777-783. (Abstract) (Html) (Pdf) [ Links ]
34. Welt CK, Schneyer AL, Heist K, Mantzoros CS. Leptin and soluble leptin receptor in follicular fluid. J Assist Reprod Genet 2003; 20:495-501. (Abstract) (Pdf) [ Links ]
35. Williams GL, Guzman Vega G, Zochi C, Gazal O. Mechanisms regulating suckling-mediated anovulation in cows. In: Stone GM, Evans G (eds). Animal Reproduction: research and practice. Anim Reprod Sci 1996; Suppl 42:289-297. [ Links ]
36. Williams GL, Mcvey WR, Hunters JF. Mammary somatosensory pathways are not required for suckling-mediated inhibition of luteinizing hormone secretion and delay of ovulation in cows. Biol Reprod 1993; 49:1328-1337. (Abstract) (Pdf) [ Links ]
37. Wiltbank MC, Gümen A, Sartori R. Physiological classification of anovulatory conditions in cattle. Theriogenology 2002; 57:21-52. (Abstract) [ Links ]
38. Wright IA, Rhind SM, White TK. A note on the note of pattern of food intake and body condition and the duration of the post-partum anoestrous period and LH profiles in beef cows. Anim Prod 1992; 54:143-146. [ Links ]
39. Yu WH, Kimura M, Walczewska A, Karanth S, McCann SM. Role of leptin in hypothalamic–pituitary function. Proc Natl Acad Sci 1997; 94:1023-1028. (Abstract) (Html) (Pdf) [ Links ]
40. Zhang Y, Proenca R, Maffei M, Barone M, Leopold L, et al. Positional cloning of the mouse obese gene and its human homologue. Nature 1994; 372:425-432. (Abstract) (Pdf) [ Links ]
¶ Para citar este artículo: Giraldo CA, Olivera Ángel M, Ruiz Cortés ZT. Efecto de la variación de peso, la condición corporal, y la expresión de receptores de leptina y de hormona luteinizante, en la anovulación posparto en vacas cebú (Bos indicus). Rev Colomb Cienc Pecu 2008; 21:228-238.
* Autor para el envío de correspondencia y la solicitud de separatas: Grupo Reproducción – Fisiología y Biotecnología, Facultad de Ciencias Agrarias, Universidad de Antioquia, AA 1226, Medellín, Colombia. Tel (+574) 2199148. E-mail: cargiraldo@gmail.com
]]>