










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Biotecnología
Print version ISSN 0123-3475
Rev. colomb. biotecnol vol.11 no.1 Bogotá Jan./June 2009
ARTÍCULO DE INVESTIGACIÓN
Inmovilización de hongos ligninolíticos para la remoción del colorante negro reactivo 5
Immobilising lignilolytic fungus for removing reactive black 5 dye
Jorge Andrés Fernández1 , Lina María Henao2 , Aura Marina Pedroza-Rodríguez3 , Balkys Quevedo-Hidalgo4
1 Microbiólogo Industrial. Pontificia Universidad Javeriana. Dirección: Carrera 71D No 122-72 Bogotá. jf2014@yahoo.es
2 Microbióloga Industrial. Dirección. Carrera 22 No 101-72. Bogotá. Correo: linahenao@hotmail.com
3Bacterióloga. Ph.D. Grupo de Biotecnología Ambiental e Industrial. Pontificia Universidad Javeriana, Carrera 7 No 43-82. Edificio Carlos Ortiz. Oficina 608, apedroza@javeriana.edu.co
4 Ingeniera Química. Ms. IQ. Grupo de Biotecnología Ambiental e Industrial. Pontifícia Universidad Javeriana, Carrera 7 No 43- 82. Edificio Carlos Ortiz. Oficina 608, bquevedo@javeriana.edu.co
Recibido: febrero 9 de 2009 Aprobado: junio 4 de 2009
Resumen
La presencia de colorantes azoicos en aguas residuales de la industria textil es un problema ambiental y sanitario, porque muchos de estos compuestos son cancerígenos. Los tratamientos biológicos son una alternativa para la remoción de ese tipo de colorantes. En el presente trabajo se evaluó el efecto de tres hongos de podredumbre blanca, Trametes versicolor, Pleurotus ostreatus y Phanerochaete chrysosporium sobre la decoloración de un agua que contiene colorante negro reactivo 5 (NR5), ampliamente usado en la industria textil. Se estudió la inmovilización de estos hongos en dos soportes, espuma de poliuretano y estropajo (L. cylíndrica) para seleccionar el mejor soporte y el hongo con mayor capacidad para la decoloración. Ambos soportes fueron igualmente efectivos, pero se seleccionó estropajo por ser un producto natural. El hongo que generó los mayores porcentajes de decoloración en 4 días fue Trametes versicolor, con 96%, 98% y 98% para agua con concentración de NR5 300 ppm, 150 ppm y 75 ppm, respectivamente. La actividad lacasa para cada concentración de NR5 fue 8 U L-1, 7 U L-1 y 5 U L-1.
Palabras clave: Trametes versicolor, Pleurotus ostreatus y Phanerochaete chrysosporium Luffa cylindrinca, espuma de poliuretano y negro reactivo 5.
Abstract
Waste water from the textile industry represents a major environmental and health problem because it contains azo dyes whose carcirogenic effect has been tested in research. Biological treatment represents a valuable alternative for removing these dyes. The effect of Trametes versicolor, Pleurotus ostreatus and Phanerochaete chrysosporium rot fungi on decoloration of water containing reactive black five (NR5) textile dye was evaluated in this work. Immobilising the fungi on polyurethane foam and luffa sponge (Luffa cylindrica) supports was studied in order to select the best support and the fungi having the best decolorisation. Both supports were equally effective; however, the luffa sponge was selected as being a natural product. Trametes versicolor produced the highest decolorisation percentages in four days (96%, 98% and 98% for 300 ppm, 150 ppm and 75 ppm NR5 concentrations, respectively) while lacase enzyme activity was 8 UL-1, 7 UL-1 and 5 UL-1 for each of them.
Key words: Trametes versicolor, Pleurotus ostreatus, Phanerochaete chrysosporium Luffa cylindrinca, polyurethane foam, reactive black 5 dye.
Introducción
Durante el proceso textil, la etapa de coloración (Dyebath), ocasiona la pérdida de considerables cantidades de colorantes. Los volúmenes liberados dependen del tipo de colorante y del proceso de aplicación, variando desde solo un 2% si se trata de un colorante básico a un 50% cuando se utilizan algunos colorantes reactivos (McMullan et ál., 2001; Levin et ál., 2004).
El color es el principal contaminante en ser reconocido en el agua, concentraciones menores a una ppm son altamente visibles para algunos colorantes, afectando así la estética de los cuerpos acuáticos (Banat et ál., 1996). Sus características químicas, los hacen muy resistentes a la luz, temperatura, al ataque microbiano y por consiguiente son compuestos altamente recalcitrantes (Levin et ál., 2004; Blánquez et ál., 2004; Romero et ál., 2006). Uno de los principales efectos adversos de su liberación al ecosistema acuático, está asociado con la reducción de la concentración del oxígeno disuelto, generando condiciones anóxicas, que pueden afectar organismos a diferentes niveles tróficos (Chander y Arora, 2007; Nilsson et ál., 2006).
Para el tratamiento de colorantes sintéticos se utilizan diversas técnicas físicas y químicas, como coagulación y floculación combinadas con filtración y flotación, precipitación-floculación con Fe(II)/Ca(OH)2, oxidación con ozono, filtración por membrana, ósmosis inversa, entre otras, las cuales producen efluentes de buena calidad. No obstante, la mayoría de estas técnicas son costosas, razón por la cual se buscan otras alternativas de tratamiento, como la utilización de hongos de podredumbre blanca como Trametes versicolor, Phanerochaete chrysosporium y Pleurotus ostreatus, que son considerados métodos biológicos no convencionales de alta eficiencia (Rodríguez et ál., 2002; Blánquez et ál., 2004; Radha et ál., 2005).
Estos microorganismos son usados para el tratamiento de efluentes textileros debido a que producen enzimas inespecíficas asociadas con la degradación de lignina, que oxidan los diferentes sustituyentes y los anillos aromáticos de un colorante. Gracias a que algunos colorantes tienen similitud estructural con las subunidades de fenilpropano que conforman el polímero de lignina, se logra que sean biotransformados o mineralizados. Las enzimas lacasa (E.C. 1.10.3.2), manganeso Peroxidasa (MnP) (E.C. 1.11.1.13), lignino Peroxidasa (E.C. 1.11.1.14) (LiP) y versátil Peroxidasa (VP) (E.C. 1.11.1.16) son las más relacionadas con este proceso; sin embargo, otras enzimas producidas por estos hongos, como cresolasas (E.C. 1.14.18.1), catecol oxidasas, glioxal oxidasas, Aril-alcohol oxidasas (E.C. 1.1.3.7) y tirosinasas, están implicadas en la degradación de sustratos más sencillos y con la producción de radicales catiónicos libres, que pueden continuar el proceso de degradación por vía enzimática o sufrir ataques nucleofílicos, para formar intermediarios menos complejos o, por el contrario se da una mineralización completa hasta CO2, agua, iones y ácidos orgánicos dependiendo del tipo de compuesto (Fernández y Henao, 2007).
La producción de biomasa es otro de los mecanismos involucrados en la remoción de colorantes y se relaciona con la capacidad de adsorción de la pared celular, compuesta en un 70% por polisacáridos cargados negativamente como la quitina y el quitosano, a los que se pueden unir por intercambio iónico una gran variedad de colorantes. Por otro lado, si la biomasa fúngica se inmoviliza en soportes biodegradables y porosos, se desarrollan portadores bifásicos de gran eficiencia, que combinan las propiedades físicas de la matriz sólida con el potencial bioquímico del hongo inmovilizado (Gómez et ál., 2008).
El objetivo de este estudio fue evaluar la capacidad de decoloración de los hongos T. versicolor, P. ostreatus y P. chrysosporium, inmovilizándolos sobre estropajo (Luffa cilíndrica) y espuma de poliuretano para degradar el colorante azóico Negro Reactivo 5 (NR5). La cepa con mayor tolerancia y potencial de decoloración fue valorada en reactor para establecer el comportamiento de enzimas ligninolíticas implicadas en la biotransformación de colorante en función del tiempo.
Materiales y métodos
Microorganismos
Se utilizaron tres cepas, T. versicolor, P. ostreatus (Pontificia Universidad Javeriana. Colombia) y P. chrysosporium (CINVESTAV-IPN México), las cuales fueron reactivadas por siembra en agar extracto de salvado de trigo: g L-1 (glucosa 10, peptona 5, extracto de levadura 2, KH2PO4 0,1, MgSO4*7H2O 0,05, MnSO4*H2O 0.076, agaragar 15, extracto líquido de salvado de trigo 175 g L-1 a 30° C por 8 días) (Pedroza et ál., 2007).
Selección del método de inmovilización para la biomasa fúngica
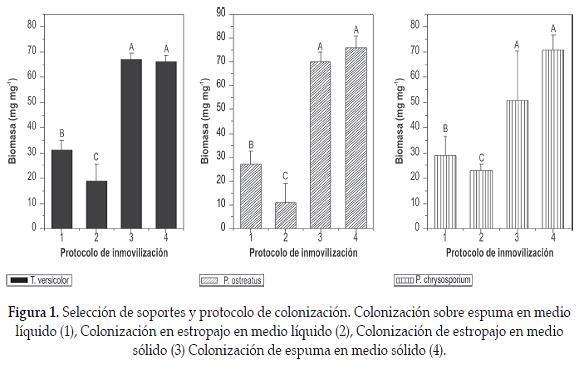
Se evaluaron dos protocolos de colonización denominados colonización en medio sólido (CMS) y colonización en medio líquido (CML), empleando como soportes estropajo (L. cylíndrica) y espuma de poliuretano de 0,5 cm3 para los dos materiales. En el sistema líquido se utilizaron erlenmeyers de 100 mL con 50 mL de caldo extracto de salvado de trigo y 5 cubos de cada tipo de soporte. Después del proceso de esterilización se inocularon con 15 discos de agar crecidos con cada hongo y se incubaron por 8 días a 30 °C y 150 rpm. El producto de inmovilización se filtró y se determinó la cantidad de biomasa atrapada en cada uno de los cubos por medio de peso seco, teniendo en cuenta el peso inicial del soporte. Este procedimiento se realizó por triplicado (Pedroza et ál., 2007).
La colonización en medio sólido se realizó colocando 30 cubos de cada material sobre una caja de Petri que contenía agar extracto de salvado de trigo y se sembraron 15 discos de los microorganismos de tal forma que estuvieran en contacto con el medio de cultivo y el soporte. Las cajas se incubaron por 8 días a 30 °C. Finalizado el proceso se determinó la cantidad de biomasa atrapada en cada cubo, por medio de peso seco. Como variable de respuesta se analizó la cantidad de biomasa fúngica atrapada, expresada como mg de biomasa/ mg de sporte. La selección del mejor soporte y método de inmovilización se realizó por medio de comparación de medias empleando el programa estadístico SAS versión 9.0® para Windows.
Selección de la cepa con mayor capacidad de decoloración a escala de erlenmeyer
Se realizó por medio de un diseño factorial 32. Las pruebas se hicieron en erlenmeyers de 100 mL que contenían 50 mL de medio Radha modificado cuya composición fue (g L-1): D-glucosa, 2,0; KH2PO4, 2,0; NH4Cl, 0,050; MgSO4*7H2O 0,5; CaCl2*2H2O, 0,1, y tiamina, 100 µg L-1. A este medio se le adicionaron 10 mL de una solución de elementos traza, cuya composición es: (g L-1): MnSO4, 0,5; FeSO4*7H2O, 0,1; ZnSO4*7H2O, 0,1. El pH final de medio fue 4,5 (Radha et ál., 2005). Cada erlenmeyer se inoculó con 5 cubos del mejor soporte colonizados con cada uno de los microorganismos que se iban a evaluar, cultivándolos por 4 días a 150 rpm. El diseño generó una matriz de 9 tratamientos realizados por triplicado, los factores estudiados fueron microorganismos y concentración de colorante en ppm. Para evaluar el efecto de los factores sobre las variables de respuesta se realizó un análisis de varianza con el programa estadístico Desing expert 6.0 ® (Montgomery, 2003).
Como variables dependientes se cuantificaron las unidades de color siguiendo el método reportado por Livernoche et ál (1983), la concentración de colorante se calculó reemplazando las absorbancias obtenidas a 597 nm en una curva de calibración de NR5 con un intervalo de 0,1-20 ppm (Fernández y Henao, 2007), las actividades enzimáticas se evaluaron siguiendo el protocolo descrito por Tinoco et ál (2001) para actividad lacasa, Michel et ál. (1991) para actividad MnP, y Tien y Kirk (1981) para lignina peroxidasa.
Selección de las condiciones de operación para el reactor biológico
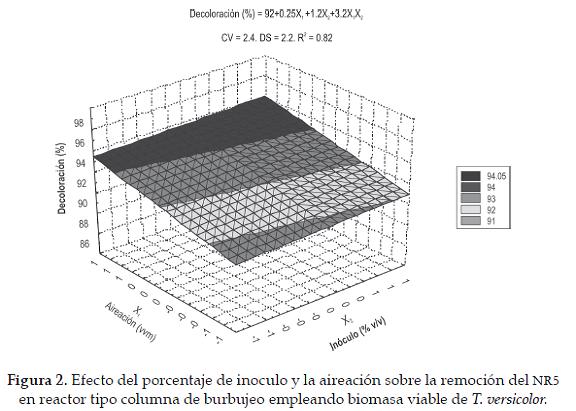
Para evaluar el efecto de dos condiciones de operación en reactor tipo columna de burbujeo sobre la capacidad de decoloración y su correlación con la actividad enzimática se llevó a cabo un diseño experimental 22. Las pruebas se realizaron en columnas de vidrio de 50 cm de largo y 5 cm de diámetro interno, que contenían un difusor de vidrio poroso (diámetro 3 mm). Cada columna fue inoculada con las diferentes concentraciones de inóculo y 500 mL del medio colorante con una concentración final de 300 ppm. El diseño generó una matriz de 4 tratamientos realizados por duplicado, los factores estudiados fueron (X1) aireación, 500 y 1000 cm3 min-1 y (X2) cantidad de inóculo (g) 5 y 9 gramos de biomasa inmovilizada y expresada como peso húmedo. El análisis de datos se realizó usando los programas SAS 9.0 ® y Desing Expert 6.0®. Los resultados fueron tratados con un modelo empírico el cual relaciona las respuestas cuantificadas con los factores evaluados y sus respectivos niveles. Para un diseño de dos factores el modelo de primer orden fue:
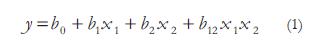
Donde y, corresponde a la variable dependiente; bo es el intercepto, b1, b2, son los coeficientes lineales y X1 X2 son los factores o variables independientes. Las variables dependientes fueron decoloración (%) y actividad lacasa (U L-1).
Curvas de remoción en reactor a escala de laboratorio
A partir de las condiciones seleccionadas en el factorial 22 se realizaron curvas de remoción por 7 días a 30 °C, realizando muestreos diarios para medir decoloración, enzimas, consumo de glucosa (Miller, 1959), y pH. Se realizaron por duplicado y como control se utilizaron cubos de soporte sin biomasa. Como análisis estadístico se llevó a cabo una comparación de medias entre tratamientos y análisis de correlación entre variables Programa SAS ® versión 9.0. Con el fin de evaluar posibles cambios estructurales en la molécula del colorante se realizaron los espectros de absorción usando un equipo de espectrofotometría Shimadzu UV/VIS 160 A. Las muestras analizadas correspondieron a los tiempos 0, 24, 48, 72 y 96 horas. Las absorbancias fueron medidas en un rango de longitud de onda entre 200 nm y 800 nm (Julson, 2005). Finalmente las características microscópicas del soporte sin colonizar y colonizado que se utilizaron en las curvas de remoción fueron observadas por microscopía electrónica de barrido, siguiendo el procedimiento descrito por Morales et ál. (2008).
Resultados
Selección del protocolo para la inmovilización de biomasa fúngica
Con respecto al protocolo de inmovilización para las tres cepas evaluadas empleando estropajo (L. cilíndrica) y espuma de poliuretano se presentaron diferencias altamente signifi - cativas entre la colonización en medio líquido (CML) y en medio sólido (CMS) (p<0,0001), demostrando que en el proceso de inmovilización en sólido se retuvo la mayor cantidad de biomasa en mg/mg de soporte, con valores de 67 mg mg-1 ± 2,3; 70 mg mg-1 ± 4 y 51 mg mg-1 ± 19 para T. versicolor, P. ostreatus y P. chrysosporiun respectivamente a los 8 días de afi anzamiento a 30 °C, sobre estropajo. Para la espuma los valores obtenidos fueron de 66 mg mg-1 ± 2,8 76 mg mg-1 ± 5 y 71 mg mg-1 5,9 para las tres cepas respectivamente (figura 1). Una vez se determinó cuál era el mejor método para la inmovilización se seleccionó un solo soporte teniendo en cuenta la cantidad de biomasa inmovilizada. De acuerdo con los resultados del análisis estadístico no se observaron diferencias significativas entre los mg de biomasa retenida/mg de espuma o estropajo (p = 0,125). Por lo tanto, cualquiera de los dos soportes podría ser empleado para el desarrollo del bioportador. Sin embargo, se seleccionó el estropajo (L. cylíndrica) por sus características de biodegradabilidad, alto contenido de lignina, celulosa y hemicelulosa que podrían favorecer la remoción de colorante.
Selección de la cepa con mayor capacidad de remoción a escala de erlenmeyer
De acuerdo con los resultados del diseño factorial 32 la interacción de X1 y X2 fue altamente signifi cativa sobre el porcentaje de decoloración (p<0,0001), demostrando que para el nivel alto de X1 (T. versicolor) los porcentajes de decoloración en los tres niveles de X2 (concentración de NR5 ppm) fueron los más elevados con 96, 98 y 98% para 300, 150 y 75 ppm del colorante (tabla 1); por lo tanto, la cepa seleccionada fue T. versicolor por tener excelente capacidad de decoloración y tolerancia a concentraciones variables del colorante. Por otra parte, P. ostreatus (nivel medio de X1), fue la segunda cepa más efi ciente para la decoloración, no obstante, las concentraciones elevadas del NR5 ejercieron un efecto negativo en la remoción con 44% a 300 ppm, lo que restringiría su uso a concentraciones de 150 y 75 ppm, en las cuales la decoloración fue del 90 y 95%. P. chrysosporium fue la cepa con menor capacidad de decoloración, con porcentajes de 16, 28 y 6% para 300, 150 y 75 ppm respectivamente.
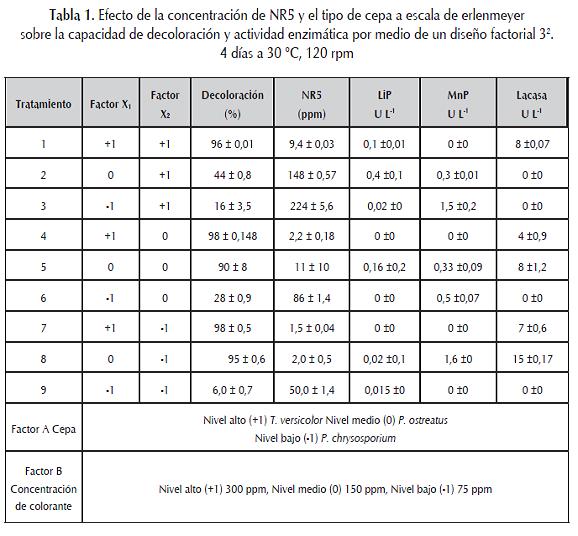
La actividad lacasa producida por T. versicolor se cuantificó al cuarto día, arrojando valores de 8 U L-1, 7 U L-1 y 5 U L-1 para 300, 150 y 75 ppm de NR5 respectivamente. Se presentó una correlación positiva significativa (p= 0,010) entre decoloración y actividad lacasa. La actividad LiP se cuantificó solamente a 300 ppm con 0,1 U L-1; resultado interesante si se tiene en cuenta que para T. versicolor la decoloración se asocia más con la enzima lacasa y es inusual que se reporte LiP.
P. ostreatus presentó actividad lacasa de 15 U L-1 y 8 U L-1 a 75 y 150 ppm. Con ausencia a 300 ppm. Las actividades MnP y LiP se cuantificaron en todos los tratamientos, representando los valores más altos del diseño. P. chrysosporium no tuvo actividad lacasa en ninguno de los tratamientos y las ligninasas dependientes de H2O2 tuvieron actividad en todos los tratamientos siendo la MnP la más elevada (tabla 1).
Condiciones de operación del reactor biológico tipo columna de burbujeo
Los resultados del análisis estadístico mostraron que la interacción de X1 y X2 en sus niveles altos fue significativa para porcentaje de decoloración y actividad lacasa (p= 0,014 y p= 0,0016). Por lo tanto, las variables dependientes se predijeron por las ecuaciones de primer orden (ecuación 2 y 3). Bajo estas condiciones cuando el reactor se inoculó con 9 g de biomasa y se aireó con 1000 cm3 min-1 se obtuvo una decoloración del 97%, que equivale a 292 ppm del colorante; con una actividad lacasa de 3.7 U L-1. La figura 2 representa los resultados del análisis estadístico ya que la mayor intensidad de color e inclinación de la superficie se presenta en la interacción de los niveles altos de cada factor.
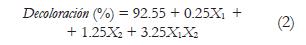
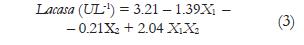
Curvas de remoción en reactor a escala de laboratorio
La decoloración fue incrementando gradualmente alcanzando un valor máximo del 98% al 5 día (294 ppm), este porcentaje se mantuvo constante hasta el final de la curva sin que se presentara un proceso de desorción del colorante. Con respecto a la actividad lacasa se observó una inducción en la actividad desde las primeras 24 horas con valores de 2 U L-1 hasta el tercer día con 8 U L-1. Posteriormente se presentó un descenso sin que se cuantificara en los días 6 y 7. La actividad lacasa y el porcentaje de decoloración se correlacionaron positivamente (p =0,048) hasta el tercer día. En cuanto al soporte sin colonizar, la decoloración fue del 31% para el primer día, alcanzando un máximo del 53% a los 7 días de proceso (figura 3).
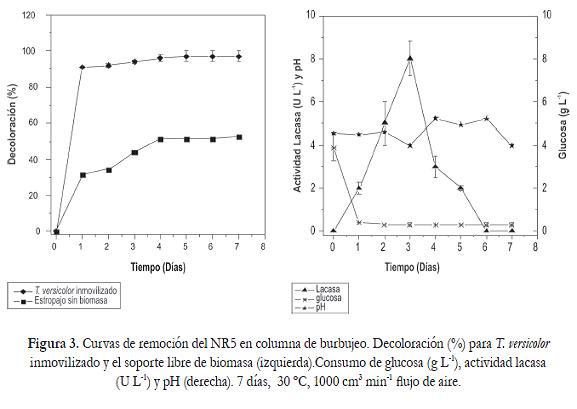
El pH se mantuvo dentro del rango de acidez, oscilando entre 3,5 y 5,2 lo que favoreció la actividad enzimática (figura 3). Por esta razón, el pH presentó una correlación positiva altamente significativa (p<0,0001) con la enzima lacasa y el consumo de glucosa. Al final del proceso la concentración residual de glucosa fue de 0,23 g L-1 y demostró que T. versicolor utilizó la fuente de carbono como co-sustrato para mantener metabolismo primario y poder llevar a cabo la biotransformación del colorante a medida que la enzima lacasa se expresa en respuesta a la inducción por los anillos aromáticos del NR5.
En el tratamiento sin biomasa la concentración de glucosa se mantuvo constante durante todo el proceso lo que permitió evidenciar que el efecto de la flora acompañante sobre la decoloración fue mínimo y el colorante fue removido por adsorción. Adicionalmente la comparación de medias entre sistema abiótico y biótico permitió establecer que existían diferencias significativas (p < 0,0001) entre tratamientos, siendo el sistema bifásico conformado por biomasa de T. versicolor y estropajo más eficiente que el soporte solo en la remoción del NR5.
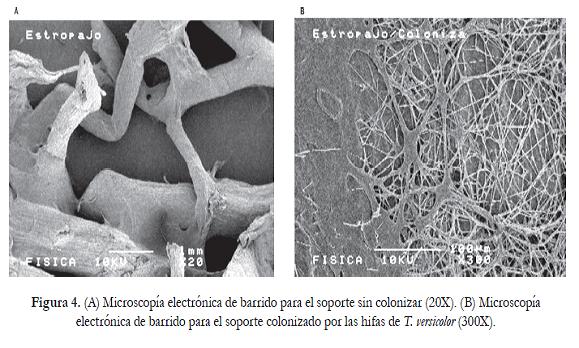
En la figura 4 se observan las microscopías electrónicas de barrido para el soporte sin colonizar y colonizado. El soporte es una matriz fi brosa, ramifi cada y porosa que favoreció la colonización de T. versicolor. En el soporte colonizado se observan dos zonas, la primera está colonizada con hifas delgadas del hongo y en la segunda se visualiza una sección superfi cial la cual está sufriendo una transformación parcial, posiblemente asociada con degradación de la lignina.
Análisis de espectros de absorción del tratamiento biológico
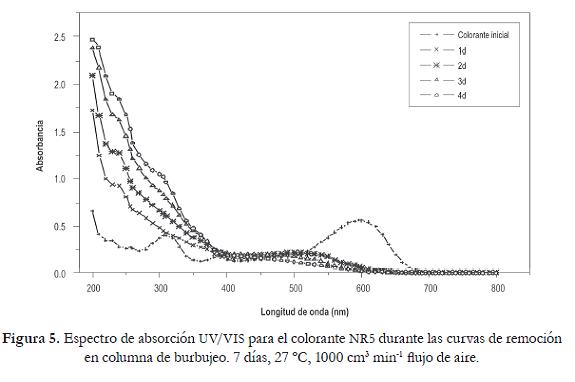
Los espectros de absorción en los diferentes días de tratamiento, muestran un desplazamiento de la longitud de onda de máxima sensibilidad del colorante (sin tratamiento) de 597 nm a 526 nm, en el primer día, lo que permite suponer que durante este tiempo se presentó un cambio en la estructura original, asociado con la transformación enzimática. El desplazamiento de la longitud de onda se hace más evidente en el trancurso de la cinética ya que al segundo día el pico de absorción se registró a 512 nm y desaparece completamente a las 72 horas (figura 5). Adicionalmente, se generaron varios picos en la región ultravioleta que se asociaron con productos aromáticos liberados por la posible ruptura del enlace azo.
Discusión
Selección del método para la inmovilización de biomasa fúngica
En un proceso de inmovilización se busca retener dentro y alrededor del material de inmovilización la biomasa para garantizar que el bioportador sea efi ciente en la remoción de contaminantes. La inmovilización en medio sólido (CMS) permitió cumplir con este objetivo, arrojando mejores resultados que en el medio líquido (CML). La posible explicación es que en el medio líquido no se recrearon las condiciones naturales de estos microorganismos, los cuales crecen colonizando la madera desde el exterior hacia el interior por medio de un proceso erosivo sobre la lignina dado por la producción de enzimas y la invasión de las hifas, bajo unas condiciones de humedad que pueden variar entre un 20 y 45% dependiendo del tipo de árbol maderable (Smith, 1998). En el sistema líquido y agitado el contenido de humedad fue superior, y el movimiento rotatorio de los erlenmeyers pudo impedir la adsorción de mayores cantidades de biomasa (Gowthman et ál., 1995; Rodríguez et ál., 2002). Por el contrario, en el cultivo estático se favoreció la inmovilización de la biomasa a los dos soportes, porque se pudo llevar a cabo la adsorción de las hifas al soporte y posteriormente estas fueron extendiéndose a través de la extensa red de poros presentes en la espuma y el estropajo.
Con respecto a la selección entre soporte inerte y no inerte no se presentaron diferencias signifi cativas entre los dos. Sin embargo, se seleccionó el soporte no inerte porque es un material rico en celulosa, hemicelulosa y lignina a partir del cual los hongos podrían obtener monómeros de fácil asimilación gracias a la acción combinada de dos grupos enzimáticos: las glicosilhidrolasas como celulasas, xilanasas y hemicelulasas, las cuales hidrolizan los enlaces glicosídicos para liberar disacáridos y monómeros como celobiosa, glucosa, xilosa, entre otros (Hou et ál.,, 2004; Couto et ál., 2005; Tanobe et ál., 2005). El segundo grupo corresponde a las polifenol oxidasas y peroxidasas que son las responsables de atacar las subunidades de fenil propano unidas por enlace C-C, éter, aril-éter y aril-aril (Moreira et ál., 2003).
El estropajo, como soporte no inerte, presenta mayores ventajas que la espuma, ya que por sus características de baja toxicidad, maleabilidad, porosidad y economía lo convierten en una excelente alternativa para el desarrollo de biodsorbentes doble propósito (Ganguly et ál., 2007). Por otro lado, la posibilidad de que los hongos obtengan a partir del soporte una fuente adicional de carbono favorece la factibilidad del procesos por que elimina o se disminuye la necesidad de suplementar el agua residual con fuente de carbono o co-sustrato para simular las condiciones ligninolíticas que favorecen la remoción de los colorantes (Moreno et ál., 2008). Al final del tratamiento biológico el subproducto sólido de la transformación del colorante o del agua residual puede ser dispuesto al incorporarlo como material llenantes en procesos de compostaje o biopilas, sin que se genere un impacto ambiental fuerte a diferencia de lo que ocurre cuando se trabaja con materiales sintéticos como espuma, anillos plásticos y poliestirenos (Liu et ál., 1998).
Por el contrario, la espuma de poliuretano es un polímero sintético y termoestable sintetizado por la mezcla del poliol y el isocinato, los cuales son compuestos químicos que no son degradados por estos hongos y simplemente son empleados como matriz porosa para facilitar la extensión apical de las hifas hasta llegar a tener contacto con los nutrientes presentes en el agar extracto de salvado de trigo.
Selección de la cepa con mayor capacidad de remoción a escala de erlenmeyer
T. versicolor fue seleccionado como microorganismo modelo para continuar con el estudio por su elevada capacidad de decoloración a diferentes concentraciones del colorante lo cual resulta muy favorable ya que los efluentes provenientes del proceso textil usualmente contienen concentraciones elevadas de estos contaminantes. Por tanto, al implementar un sistema biológico es necesario que la cepa tolere estas concentraciones para garantizar que la remoción de color y disminución en la toxicidad se lleve a cabo por medio de mecanismos bioquímicos y físicos (Rodríguez et ál., 2002; Zille et ál., 2003; Romero et ál., 2006).
P. ostreatus también fue una cepa eficiente para la decoloración, como lo han reportado Rodríguez et al. (1999). No obstante, las concentraciones elevadas del colorante ejercieron un efecto negativo en la remoción (44%), limitando su uso a valores menores. Lo anterior está asociado quizás a algún tipo de inhibición relacionada con los productos intermediarios de la degradación a altas concentraciones de colorante, que podrían estar afectando al hongo directamente o a la producción de las enzimas implicadas en el proceso. Esto ocasionaría el empleo de una dilución previa del agua para disminuir la concentración del colorante a niveles aptos para ser soportados por P. ostreatus, lo cual implicaría gastos operacionales adicionales. Por otro lado, P. chrysosporium fue la cepa con menor capacidad de decoloración, por lo tanto, para este tipo de colorantes no sería una cepa muy recomendable.
Con respecto a la actvidad lacasa producida por T. versicolor en las tres concentraciones se pudo demostrar que es una de las enzimas implicadas en la biotransformación del NR5. Según Zille et ál. (2003) la lacasa reacciona con el NR5 al darse una interacción electrostática entre las fracciones aniónicas del colorante (Grupos sulfónicos) y los aminoácidos cargados de la enzima, la interacción se ve favorecida por la presencia de ciertas sales como cloruro de sodio, sulfato de amonio, etc, las cuales incrementan la fuerza iónica de la solución. Al analizar la correlación entre actividad lacasa y decoloración los resultados obtenidos en este estudio coinciden con los trabajos realizados por Rodríguez et ál. (1999) y Zille et al. (2005), quienes observaron que la lacasa está implicada en la degradación de colorantes azo y más específicamente en la decoloración del NR5 (Zille et ál., 2003). Aunque la actividad lacasa en T. versicolor a 75 ppm fue más baja (8 U L-1) si se compara con la de P. ostreatus a la misma concentración (15 U L-1), diferentes estudios han demostrado que incluso la lacasa producida por la misma especie de hongo, o por especies diferentes, puede tener afinidad por sustratos distintos, lo que podría afectar de manera relevante el grado de degradación de un sustrato (Mayer y Staples, 2002). A pesar de que P. ostreatus presentó la mayor actividad lacasa, puede tener un grado de afinidad menor por el colorante, en comparación con la enzima producida por T. versicolor.
Condiciones de operación del reactor biológico tipo columna de burbujeo
Los resultados del análisis estadístico mostraron que la interacción de X1 y X2 en sus niveles altos fue significativa. Se obtuvieron los mejores resultados debido a que la transferencia de oxígeno es más eficiente que a nivel de erlenmeyer (Pedroza et ál., 2007). Sin embargo, la baja actividad enzimática en el tratamiento que presentó la mayor decoloración podría explicarse por la acción de otro tipo de enzimas, no cuantificadas, como las tirosinasas (E.C. 1.14.18.1), de las que se ha encontrado relación con la degradación de azo colorantes (Jadavh et ál., 2007). Estas enzimas pueden actuar de manera indirecta en la degradación del colorante, atacando compuestos de bajo peso molecular menos complejos que la molécula original previamente atacada por las enzimas de mayor poder oxidativo (LiP, MnP y lacasa), o mediante la producción de compuestos de alto poder oxidante como el peroxido de hidrógeno, asociado con la actividad de las glioxal oxidasas (Ander y Marzullo, 1997). Por otra parte, aunque las enzimas implicadas en la degradación de la lignina son de carácter extracelular, la medición de la lacasa intracelular ha sido contemplada en la metodología de algunos estudios, ya que está implicada con la degradación de colorantes (Blánquez et ál., 2004). Romero et ál. (2006) proponen que en el primer paso en la degradación de colorantes azo hay una adsorción de estos y su posterior transferencia al interior de la célula fúngica, en donde sus mecanismos de degradación intracelulares entrarían en acción por lo que la expresión de bajas concentraciones de enzimas de tipo extracelular en ocasiones no está relacionada con altos porcentajes de degradación de un compuesto problema.
Curvas de remoción
Al analizar la tendencia de la decoloración en función del tiempo el incremento gradual y elevado obtenido desde las primeras horas de proceso es similar a lo reportado por Blánquez et al. (2004). En su estudio obtuvieron 83% de decoloración aproximadamente durante el mismo periodo de tiempo. Así mismo, Borchert y Libra (2001) demostraron que el 65% del NR5 fue removido a las 24 horas. En el presente estudio se pudo presentar un sinergismo entre la adsorción y la presencia de enzimas, ya que inicialmente el colorante se une por medio de enlaces covalentes tanto al soporte como a la biomasa inmovilizada, proceso que se realiza con rapidez, permitiendo que los grupos cromóforos quedaran cercanos a las enzimas ligninolíticas presentes en la superficie de las hifas tal y como lo demostraron Evans et ál. (1991) y Yang et ál. (2003). Posteriormente, las enzimas empezarían a realizar la biotransformación del colorante, generando una disminución en la concentración y cambios en los espectros de absorción en la región visible. Adicionalmente, la decoloración también fue atribuida a la unión del NR5 al soporte, ya que este tipo de colorante pertenece a una familia de los reactivos y están diseñados específicamente para adherirse fuertemente a las fibras celulósicas como el algodón, por esta razón el estropajo solo removió el 53% al día 7. Este resultado es esperado si se tiene en cuenta que la pérdida de algunos colorantes reactivos en los procesos de teñido es de aproximadamente el 50% (McMullan et ál., 2001; Levin et ál., 2004).
Con respecto a la actividad lacasa la producción se fue incrementando desde las 24 horas con un máximo al tercer día, posiblemente porque la estructura aromática del colorante y el soporte pudieron realizar un proceso de inducción enzimática (Rodríguez el ál., 2002; Amaral et ál., 2004). El evidente cambio de color en el residual durante los 7 días de tratamiento, variando entre el púrpura, rojo y amarillo, demuestran una transformación del complejo auxocromo-cromóforo, que se asocia con un ataque enzimático del grupo hidroxi de la molécula del colorante. Al desestabilizar este complejo la absorción de luz disminuye y, por tanto, el color de la sustancia cambia. Otro factor que favoreció la actividad lacasa fue que no se presentó un cambio notorio en el pH el cual se mantuvo ácido por el metabolismo de la glucosa con la respectiva producción de ácidos orgánicos como el oxálico, manteniendo el pH en valores muy cercanos al pH óptimo de la enzima tal como lo reportaron Rancaño et ál. (2003), que en sus estudios demostraron que el pH óptimo para la actividad de las enzimas ligninolíticas, se encuentra entre 4,5-5,0, con una inactivación parcial por encima de 5,5 y por debajo de 3,5.
El consumo acelerado de la glucosa durante las primeras 24 horas pudo relacionarse con el empleo de la misma como fuente de carbono sencilla para mantener el metabolismo primario mientras se llevaba cabo la remoción del colorante y al hacerse limitante el microorganismo empezaría a emplear otras fuentes disponibles como el material lignocelulósico presente en el soporte; por esta razón, en las microscopías electrónicas de barrido se observaron cambios estructurales en el estropajo con zonas de colonización abundante por parte de las hifas de T. versicolor. Un resultado similar fue presentado por Garzón et ál. (2009), quienes inmovilizaron a P. ostreatus y T. versicolor en mallas de agave tequilero y observaron por medio de microscopías electrónicas que el soporte era colonizado por la biomasa fúngica y, a medida que transcurrió el tiempo fue degradado presentándose un incremento en los miligramos de biomasa y en la cantidad de azúcares totales que fueron liberados al medio sintético suplementado con colorantes azóicos, trifenilmetánicos y antraquinónicos.
Al realizar un análisis del espectro de absorción del colorante, se pueden mostrar en cierta forma los cambios del complejo cromóforoauxocromo del colorante, al observar el desplazamiento del espectro de absorción con un marcado incremento de los picos en la región UV que se asocia a un cambio drástico de la molécula del NR5, lo que supone la biotransformación completa de éste. Borchert y Libra (2001) realizaron el mismo tipo de análisis, demostrando que al igual que en este estudio, el corrimiento de la longitud de onda de luz visible a luz UV se debe a cambios en la estructura química del colorante, y que esto evidencia la presencia de metabolitos aromáticos en el producto de degradación, lo que haría pensar en una posible toxicidad del compuesto por la presencia de aminas aromáticas. Es de destacar el color amarillo que se obtuvo al final de los tratamientos en ambos estudios.
Conclusión
De las tres cepas evaluadas se seleccionó a T. versicolor para desarrollar un bioportador a base de biomasa viable inmovilizada por colonización en sólido (CMS) sobre estropajo con elevada porosidad, adsorción y actividad biológica para la remoción del colorante azóico negro reactivo 5. Este fue eficiente a diferentes concentraciones del colorante, y bajo condiciones de operación a escala de laboratorio asociadas con elevada concentración de inóculo y aireación alta fue capaz de remover el 98% de la concentración inicial mediante un proceso combinado entre adsorción y actividad enzimática del tipo lacasa.
Agradecimientos
El presente estudio forma parte del proyecto No 00002730 registrado en la Oficina de Fomento a la Investigación de la Vicerrectoría Académica. Pontificia Universidad Javeriana. Bogotá, Colombia.
Al doctor Ciro Falcony Guajardo y a la maestra Ana Bertha Soto Guzmán del Departamento de Física del CINVESTAV-IPN. México D.F por la autorización y colaboración para el procesamiento de muestras para microscopía electrónica de barrido.
Referencias bibliografícas
1 Amaral, P. F., Fernández, D. L., Tavares, A. P., Xavier, A. B., Cammarota, M. C., Coutinho, J. A., Coelho, M. A. 2004. Decororization of dyes from textile wastewater by Trametes versicolor. Environmetal Technology 25: 1313-1320. [ Links ]
2 Ander, P., Marzullo, L. 1997. Minireview sugar oxdioreductases and veratryl alcohol oxidase as related to lignin degradation. Journal Biotechnology 53: 115-131. [ Links ]
3 Banat, I., Nigam, P., Singh, D. and Marchant, R. 1996. Microbial decolorization of textile-dye-containing effluents: A review. Bioresource Technology 58: 217-225. [ Links ]
4 Blánquez, P., Casas, N., Font, X., Gabarrell, X., Sarrá, M., Caminal, G., Vicent, T. 2004. Mechanism of textile metal dye biotransformation by Trametes versicolor. Water Research 38: 2166-2172. [ Links ]
5 Borchert, M. and Libra, J. 2001. Decolorization of reactive dyes by white rot fungus Trametes versicolor in sequencing. Biotechnology & Bioengineering. 75: 313-321. [ Links ]
6 Dhaouadi, H., Poncin, S., Hornut, J., Midouxb, N. 2007. Gas-liquid mass transfer in bubble column reactor: Analytical solution and experimental confirmation. Chemical. Engeenring Proccesing doi:10.1016/j. cep.2006.11.009. [ Links ]
7 Chander, M. and Arora, D. 2007. Evaluation of some whiterot fungi for their potential to decolourise industrial dyes. Dyes and Pigments 72: 192-198. [ Links ]
8 Chen, K., Wu, J., Liou, D. 2003. Decolorization of the textile dyes by newly isolated bacterial strains. Journal Biotechnology 101: 57-68. [ Links ]
9 Couto, S., Rosales, E., Gundin, M., Saroman, M.A. 2004. Explotation of a waste from the brewing industry for laccase production by two Trametes species. Journal of Food Engineering 64: 423-428. [ Links ]
10 Fernández-González, J. y Henao-Jaramillo, L. 2007. Hongos de la podredumbre blanca y fotocatálisis homogénea nanoestructurada con TiO2 como alternativa biotecnológica para la remoción del negro reactivo 5. Trabajo de grado. Pontificia Universidad Javeriana, Colombia. 164 pg. [ Links ]
11 Evans, C.S., Gallagher, I.M., Atkey, P.T. Wood, D.A. 1991. Localization of degradative enzymes in white-rot decay of lignocellulose. Biodegradation 2: 93. [ Links ]
12 Ganguly, R., Dwivedi, P., Singh, R. 2007. Production of lactic acid wiht luffa sponge immobilized Rhizopus oryzae RBU 2-10. Bioresource Technology 8: 1246-1251. [ Links ]
13 Garzón, C., Pedroza, A.M., Barragán, B. 2009. Biodecoloración de colorantes azóicos, trifenilmetánicos y antraquinonicos por cepas de P. ostreatus, T. versicolor y K. neumaniae. Trabajo de grado. Escuela Nacional de Ciencias biológicas del Instituto Politécnico Nacional. México. D. F. México, 150 pg. [ Links ]
14 Gómez-Bertel, S., Amaya-Bulla, D., Maldonado-Saavedra, C., Martínez-Salgado, M.M., Quevedo-Hidalgo, B., Soto-Gúzman, A., Pedroza-Rodríguez, A.M. 2008. Evaluación de Phanerochaete chrysosporium, Trametes versicolor, Pleurotus ostreatus y Aspergillus niger como alternativa para el tratamiento de aguas residuales del curtido de pieles. Revista Internacional de Contaminación Ambiental 24: 93-106. [ Links ]
15 Gowthman, K., Raghava, K., Ghildyal, N., Karanth, N. 1995. Estimation of KLA in solid state using and packed- bed bioreactor. Process Biochemistry 30: 9-15. [ Links ]
16 Hou, H., Zhou, J., Wang, J., Du, C., Yan, B. 2004. Enhancement of laccase production by Pleurotus ostreatus and its use for the decolorización of antraquinone dye. Process Biochemistry 39: 1415- 1419. [ Links ]
17 Jadahv, J., Parshetti, G., Kalme, S., Govindwar, S. 2007. Decolourization of azo dye methyl red by Sacharonmyces cerevisea MTCC463. Chemosphere 68: 394-400. [ Links ]
18 Julson, A. 2005. Photocatalytic decolorization of organic dyes in titanium dioxide-air systems. Master Thesis. Department of Chemical and Biomolecular Enineering; North Carolina State University. North Carolina. North Carolina State University, 205 pg [ Links ]
19 Levin, L., Papinutti, L., Forchiassin, F. 2004. Evaluation of Argentinean white rot fungi for their ability to produce lignin modifying enzymes and decolorize industrial dyes. Bioresource. Technology 94: 169-176. [ Links ]
20 Liu, Y., Seki, M., Tanaka, H. 1998. Characteristics of Loofa (Luffa cylindrica) sponge as a carrier for plant cell immobilization. Journal of Fermentation and Bioengineering 85: 416-421. [ Links ]
21 Livernoche, D., Jurasek, L., Desrochers, M., Dorica, J. 1983. Removal of Color form kraft Mill Wastewaters whit Cultures of White-rot Fungi and with Immobilized Mycelium of Coriolus versicolor. Biotechnology & Bioengineering 25: 2055-2065. [ Links ]
22 Mayer, A. and Staples, R. 2002. Laccase new functions for and old enzyme. Phytochemistry 60:551-565. [ Links ]
23 McMullan, G., Meehan, C., Conneely, A., Kirby, N., Robinson, T., Nigam, P., Banat, I., M, Marchant, R., Smyth, W. 2001. Microbial decolourisation and degradation of textile dyes. Applied Microbiology Biotechnology 56: 81- 87. [ Links ]
24 Michel, F., Balachandra, S., Grulke, E., Adinarayana, C. 1991. Role of manganese peroxidases and lignin peroxidases of Phanerochaete chrysosporium in the decolorization of kraft bleach plant effluent. Applied Environmental Microbiology 157: 2368-2375. [ Links ]
25 Miller, G. 1959. Use of dinitrosalicyclic acid reagent for determination of reducing sugar. Analytical Chemistry. 31: 426-428. [ Links ]
26 Montgomery. Diseño y Análisis de experimentos. 2003. Editorial Limusa. Segunda edición. 218- 276. [ Links ]
27 Morales, D.M., Ruiz, K.J., Martínez, M.M., Soto, A., Falcony, C., Pedroza, A.M. 2008. Bioadsorción de Ní, Pb y Cd sobre la pared fúngica de Phanerohaete chrysosporium para la remoción de metales pesados. En Memorias del XI Congreso Nacional de Microscopía. León Guanajuato. México. [ Links ]
28 Moreira, M., Feijoo, G., Lema, J. 2003. Fungal bioreactors: Applications to white-rot fungi. Reviews. Environmental Science and Biotechnology 2003 ; 2: 247-259. [ Links ]
29 Moreno, N., Ospina, A., Puentes, J., Pedroza, A.M. 2008. Optimización de inductores metálicos y co-sustrato en la remoción de negro reactivo 5 empleando a Pleurotus ostreatus inmovilizado en fique. En memorias de Congreso Internacional de Gestión sostenible del agua: Reutilización, tratamiento y evaluación de la calidad. Medellín. Colombia. [ Links ]
30 Murugesan, K., Dhamija, A., Nam, I., Kim, Y., Chang, Y. 2006. Decolourization of reactive black 5 by laccase: optimization by response surfface methodology. Dyes and Pigments 20: 1-9. [ Links ]
31 Nilsson, I., Möller, A., Mattiasson, B., Rubindamayugi, M. 2006. Decolorization of Synthetic and Real Textile Wastewater by the Use of White-rot Fungi. Enzyme Microbiology Technology 38: 94-100. [ Links ]
32 Pedroza, A.M., Matiz, A., Quevedo, B. 2003. Manual de laboratorio de Introducción a la Biotecnología. Bogotá. Javegraf. 116 pg. [ Links ]
33 Pedroza, A.M., Mosqueda, R., Alonso-Vante, N., Rodríguez- Vázquez, R. 2007. Secuencial treatment via T. versicolor and UV/TiO2/RuxSey to reduce contaminants in waste water resulting from the bleaching process during paper production. Chemosphere 67:793-801. [ Links ]
34 Radha, K., Regupathi, I., Arunagiri, A., Murugesan, T. 2005. Decolorization of synthetic dyes using Phanerochaete chrysosporium and their kinetics. Process Biochemistry 40: 3337-3345. [ Links ]
35 Rancaño, G., Lorenzo. M., Molares, N., Rodríguez, S., San Román, M. 2003. Production of laccase by T. versicolor in an Airlift fermentor. Process Biochemistry 39: 467-473. [ Links ]
36 Rodriguez, E., Pickard, M. and Vazquez-Duhalt, R. 1999. Industrial dye decolorization by laccases from ligninolytic. Current Microbiology 38: 27-32. [ Links ]
37 Rodríguez, S., Gundín, M., Lorenzo, M., Sanromán, A. 2002. Screening of supports and inducers for laccase production by Trametes versicolor in semi-solid-state conditions. Process Biochemistry 38: 249-255. [ Links ]
38 Romero, S., Blánquez, P., Font, X., Gabarrell. X., Sarrá, M., Caminal, G., Vicent, T. 2006. Different approaches to improving the textile dye degradation capacity of Trametes versicolor. Biochemistry Engineering Journal 6: 17-24. [ Links ]
39 Smith, W.F. 1998. Fundamentos de la Ciencia e Ingeniería de los materiales. Madrid España: McGraw-Hill. P 6711. [ Links ]
40 Tanobe, V., Sydenstricker, T., Munaro, M., Amico, S. A comprehensive characterization of chemically treated Brazilian sponge-gourds (Luffa cylindrica). 2005. Poly test 24: 474-4782. [ Links ]
41 Tien, M. and Kirk, T. 1983. Lignin degradation enzyme from the hymenomycete. Phanerochaete chrysosporium burds. In. Science. 221: 661-663. [ Links ]
42 Tinoco, R., Pickard, M. and Vazquez-Duhalt, R. 2001. Kinetic differences of purifed laccases from six Pleurotus ostreatus strains. Letters Applied Microbiology 32:331-335. [ Links ]
43 Yang, Q., Yang, M., Pritsch, K., Yediler, A., Hagn, A., Schloter, M., Kettrup, A. 2003. Decolorization of synthetic dyes a production of manganese-dependent peroxidase by new fungus isolates. Biotechnology Letters 25: 709-716. [ Links ]
44 Zille, A., Górnacka, B., Rehorek, A. and Cavaco-Paulo, A. 2005. Degradation of Azo Dyes by Trametes villosa Lacasse over Long Periods of Oxidative Conditions. Applied Environmental Microbiology 71: 6711-6718. [ Links ]
45 Zille, A., Tzankov, T., Gübitz, G., Cavaco-Paulo, A. 2003. Immobilized laccase for decolourization of Reactive Black 5 dyeing effluent. Biotechnology Letters 25:1473-1477. [ Links ]

