










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkRevista Colombiana de Entomología
versión impresa ISSN 0120-0488versión On-line ISSN 2665-4385
Rev. Colomb. Entomol. v.31 n.2 Bogotá jul./dic. 2005
Capacidad reproductiva de Supputius cincticeps (Hemiptera: Pentatomidae): se afectan las hembras con la continua exposicion al macho?
Reproductive capacity of Supputius cincticeps (Hemiptera: Pentatomidae): are females affected by continual exposure to males?
RODRIGO DINIZ SILVEIRA1, ADRIAN JOSE MOLINA-RUGAMA1, JOSE COLA ZANUNCIO1
1Departamento de Biologia Animal, Universidade Federal de Viçosa/BIOAGRO. 36.570-000, Viçosa. Minas Gerais, Brasil. Fax: 55-31-3899-4012. E-mail: ajmolina@insecta.ufv.br
Resumen. Se estudio la influencia del periodo de exposicion al macho sobre la capacidad reproductiva de las hembras deSupputius cincticeps (Stal) (Hemiptera: Pentatomidae) en laboratorio. El experimento se realizo a una temperatura de 25.0 ± 2.0ºC, humedad relativa de 60.0 ± 10.0% y fotoperiodo de 12 h. Las hembras se expusieron al macho para apareamiento, durante toda la vida (T1), hasta la primera puesta (T2) y sin exposicion (T3). Se registro el numero de puestas, el numero y la viabilidad de los huevos y la longevidad de las hembras. El numero de puestas fue semejante entre Tl y T2 (5.5 ± 0.88 y 4.9 ± 1.09 respectivamente) y mayores que T3, (1.8 ± 0.34). Igualmente, el numero y la viabilidad de los huevos de las hembras dc 5. cincticeps entre Tl y T2 fueron semejantes (61.0 ± 10.74 y 49.9 ± 11.97 huevos y 62.2 ± 3.59 y 65.1 ±4.09% respectivamente). Las chinches sin aparear (T3) colocaron menor numero de huevos por hembra (12.2 ± 2.70) y estas puestas no fueron viables. Por otro lado, la longevidad de las hembras de S. cincticeps fue semejante entre los tratamientos con valores de 28.0 ± 2.25 (Tl), 24.4 ± 3.48 (T2) y 26.9 ± 2.54 (T3) dias, lo que sugiere que esa caracteristica es independiente del apareamiento. Para cria masiva, las hembras y machos de S. cincticeps pueden mantenerse en produccion por veints a veinticinco dias, pues durante ese periodo obtuvieron mas del 80% del total de huevos producidos.
Palabras clave: Control biologico. Chinche predadora. Cria artificial. Seleccion sexual. Reproduccion.
Summary. The influence of the period of male exposure on the reproductive capacity of Supputius cincticeps (Stal) (Hemiptera: Pentatomidae) females in laboratory was studied. The experiment was conducted at a temperature of 25.0 ± 2.0"C, relative humidity 60.0 ± 10.0% and photoperiod 12 h. Females of 5. cincticeps were exposed to the male for mating during their entire lifetime (Tl), until laying the first egg mass (T2) and without male exposure (T3). The variables observed were number of egg masses, number and viability of the eggs and longevity of the female. The number of egg masses was similar between Tl and T2 (5.5 ±0.88, and 4.9 ± 1.09 per female respectively), and higher than T3 (1.8 ±0.34). The number and viability of eggs of S. cincticeps were also similar between Tl and T2, (61.0± 10.74 and 49.9 ± 11.97 eggs, and 62.2 ± 3.59% and 65.1 ±4.09%, respectively). Bugs that did not mate (T3) laid fewer eggs per female (12.5 ±2.70) and these were not viable. Longevity of females of S, cincticeps was similar among treatments with values of 28.0 ± 2.25 (Tl), 24.4 ± 3.48 (T2) and 26.9 ± 2.54 (T3) days, showing that this biological characteristic is independent of mating. In terms of mass rearing, the females and males of S. cincticeps can be maintained in production for twenty to twenty five days old because during this period the bugs achieved more than 80% of total egg production.
Key words: Biological Control. Predator bug. Artificial rearing. Sexual Selection. Reproduction.
Introduccion
Las chinches predadoras de la familia Pentatomidae, subfamilia Asopinae, son agentes importantes de control biologico que se alimentan sobre las diferentes fases de desarrollo de insectos pertenecientes a los ordenes Lepidoptera, Coleoptera, Hymenoptera y Hemiptera (Zanuncio et al. 1994; De Clercq et al. 2002). El establecimiento de estas especies en un ecosistema depende de diferentes factores bioticos y abioticos los cuales pueden afectar su reproduccion y longevidad (Molina-Rugama et al. 1997, 1998; Mourao et al. 2003). No obstante, el exito reproductivo de las especies poliandricas puede ser explicado por los apareamientos continuos, como se observa en las hembras de Podisus spp. las cuales aparentemente necesitan copular con diferentes machos para mantener una alta viabilidad de los huevos (Torres et al. 1997). De esa forma se espera que especies que copulan mas de una vez, cuando apareadas en ambientes confinados, presenten una mayor tasa de copulas y consecuentemente un aumento en el "fitness" reproductivo (Arnqvist y Nilsson 2000). Sin embargo, un experimento realizado con hembras de la araña Stegodyphus lineatus Latreille (Araneida: Eresidae) demostro que el aumento en la tasa de apareamiento redujo el exito reproductivo en un 35% y afecto la masa corporal de los descendientes (Maklakov et al. 2005).
La chinche predadora Supputius cincticeps (Stal) (Hemiptera: Pentatomidae) se caracteriza por presentar tres copulas como media (A.J. Molina-Rugama, observacion personal) dependiendo de su longevidad, pudiendolo hacer con machos diferentes o con el mismo macho cuando se mantienen en un mismo ambiente. Esta especie predadora ha sido encontrada en baja densidad en reforestaciones con eucalipto, lo que hace necesario el desarrollo de estudios sobre su biologia, reproduccion y cria masiva para que este predador pueda ser multiplicado y liberado en programas de control biologico (Zanuncio et al. 1996-1997). Actualmente, este pentatomido es alimentado con larvas de Tenebrio molitor L. (Coleoptera: Tenebrionidae); los adultos son mantenidos dentro de potes plasticos de 500 ml en una proporcion 9 : S de 1:1. En este estudio se determino el efecto del periodo de exposicion al macho sobre la fecundidad y la supervivencia de las hembras S. cincticeps.
Materiales y Metodos
El experimento se realizo en el laboratorio de Entomologia Forestal, de la Universidad Federal de Viçosa, Viçosa, Estado de Minas Gerais, Brasil. Los adultos de S. cincticeps procedentes de la cria mantenida en el referido laboratorio se colocaron en camaras de crecimiento a una temperatura de 25.0 ± 2 °C, humedad relativa de 60.0 ± 10% y fotoperiodo de 12 hs. La colonia de S. cincticeps se establecio aproximadamente hace cinco años con reintroduccion de individuos del campo por lo menos una vez al año para evitar el efecto de consanguinidad. Durante ese periodo los predadores recibieron como presas larvas o pupas de Tenebrio molitor L. (Coleoptera: Tenebrionidae).
Para verificar el efecto del confinamiento de la pareja, la hembra virgen de S. cincticeps con aproximadamente 24 h de edad se apareo con un macho de edad semejante, de acuerdo con los siguientes tratamientos: hembras expuestas al mismo macho durante toda la vida (Tl); apenas hasta la primera puesta (T2) y sin exposicion al macho (T3). Los tratamientos se definieron con base en la metodologia de cria de esos insectos en laboratorio (machos y hembras juntos hasta el final de su ciclo de vida) y al periodo de dias necesario para que las hembras coloquen su primera puesta (alrededor de seis dias) (Zanuncio et al. 2001). Los machos que murieron antes de la hembra correspondiente se substituyeron por otros provenientes de la cria con forme cada tratamiento. Las hembras utilizadas pesaron entre 50.0 y 55.0 mg para evitar el efecto de tamaño sobre las variables reproductivas estudiadas (Zanuncio et al. 2002). La pareja de S. cincticeps se mantuvo aislada y alimentada con pupas de T. molitor (Zanuncio et al. 1996) y diariamente se observo la supervivencia de los adultos y la presencia de puestas, las cuales se retiraron y se colocaron en un pedazo de algodon dentro de una placa de Petri conteniendo otro pedazo de algodon humedecido. De esa forma se determino: el numero de huevos, el numero de huevos por puesta, el numero de puestas, el porcentaje de huevos viables y la longevidad de las hembras.
El experimento se organizo bajo un diseño completamente aleatorio, con tres tratamientos y 20 repeticiones. Cada unidad experimental estaba constituida por una pareja de S. cincticeps. Los datos correspondientes al numero de huevos por puesta se transformaron en 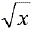 y la viabilidad y la cantidad de huevos por hembra en n1 = arcsen
y la viabilidad y la cantidad de huevos por hembra en n1 = arcsen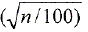 . Los promedios obtenidos se compararon por la prueba de Tukey al nivel de 5% de probabilidad (Gomes 1978).
. Los promedios obtenidos se compararon por la prueba de Tukey al nivel de 5% de probabilidad (Gomes 1978).
Resultados
El numero de huevos colocados por S. cincticeps fue afectado por los diferentes tratamientos estudiados (F = 6.12; df = 2, 39; P = 0.0048). La cantidad de huevos registrados en las hembras apareadas durante toda la vida (T1 = 61.0 ± 10.74) o hasta la primera puesta (T2 = 49.9 ± 11.97) fueron semejantes entre si y mayores a aquellos obtenidos con las hembras virgenes (T3 = 12.2 ± 2.70). La misma tendencia fue observada para el numero promedio de huevos por puesta (F = 4.89; df = 2, 17; P = 0.008) y el numero de puestas por hembra (F=4.66;df=2,39;P=0.015). Los promedios obtenidos para esas variables fueron similares entre los tratamientos T1 y T2 (11.1 ± 0.64 y 10.3 + 0.73; 5.5 ± 0.88 y 4.9 ± 1.09, respectivamente), mientras tanto, aquellas del tratamiento T3 tuvieron los menores valores con 6.7 ± 1.07 y 1.8 ± 0.34, respectivamente. La viabilidad de los huevos fue semejante entre las hembras del T1 y T2, con valores de 62.2 ± 3.59 y 65.1 ± 4.09%, respectivamente (F = 0.28; df = 1, 15; P = 0.594). No hubo eclosion de huevos en las hembras del tratamiento T3, por lo que este grupo no fue incluido en el analisis de varianza.
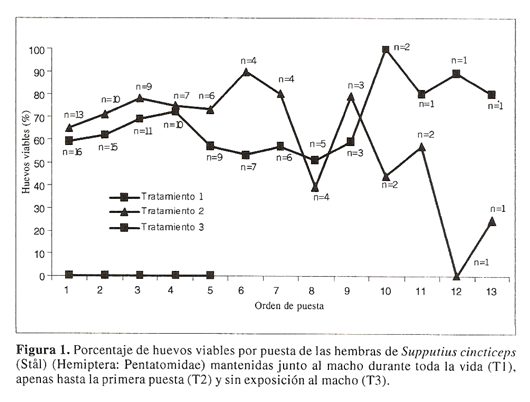
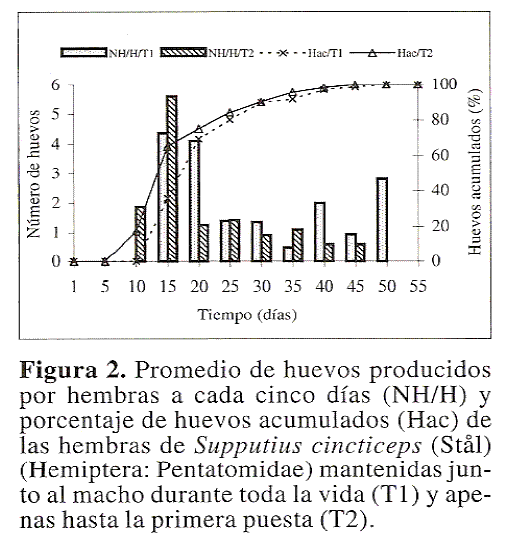
Al interpretar la curva correspondiente a la viabilidad de los huevos en funcion del orden de puesta (Fig. 1) se observa que las hembras de S. cincticeps del tratamiento T2 tuvieron una drastica reduccion en el porcentaje de huevos viables cuando son comparadas con aquellas del tratamiento T1, las cuales mantuvieron un alto porcentaje de eclosion al final de su periodo de reproduccion. De otra parte, las hembras de S. cincticeps de los tratamientos T1 y T2 comenzaron a realizar puestas entre el quinto y decimo dias, llegando a colocar 80% de los huevos en los primeros 25 dias (Fig. 2). Por otro lado, la longevidad de las hembras de S. cincticeps fue semejante entre los tratamientos (F = 0.42; df = 2,59; P = 0.655), con valores promedios de 28.0 ± 2.25, 24.4 ± 3.48 y 26.9 ± 2.54 dias para los tratamientos T1, T2 y T3, respectivamente.
Discusion
La fecundidad de las hembras de S. cincticeps no estuvo afectada por el periodo de exposicion al macho. Sin embargo, Torres et al. (1997) mencionaron que las hembras de Podisus nigrispinus (Dallas) (Hemiptera: Pentatomidae) copuladas durante toda la vida presentaron menor numero de huevos en relacion con aquellas apareadas una, dos o cuatro veces, lo que puede ser debido a los diferentes cortejos e intensidades de copula realizados por el macho. Por otro lado, el hecho de que las hembras expuestas al macho durante todo su ciclo de vida (T1) o apenas hasta la primera puesta (T2) hayan presenlado desempeño reproductivo similar, demuestra que los machos pueden mantenerse dentro del vaso de cria hasta la primera puesta y posteriormente liberarse en el campo. Mientras tanto, las hembras de S. cincticeps virgenes (T3) presentaron menor numero de puestas y de huevos, lo que tambien fue observado por De Clercq y Degheele (1997) y por Torres et al. (1997) para P. maculiventris y P. nigrispinus, respectivamente.
Aproximadamente, 20% de las hembras dc S. cincticeps no realizaron puestas, lo que representaria una reduccion de casi 244 huevos para un grupo de 20 hembras de este predador. Esto puede estar asociado a la falta de un apareamiento adecuado entre los individuos, ya sea por no ocurrir copula o inclusivo por haber resistencia de las hembras a nuevas copulas. Rodriguez (1998), estudiando el comportamiento de apareamiento de Ozophora baranowskii (Hemiptera: Lygaeidae) observo que durante la copula, las hembras resistian a la misma golpeando el abdomen del macho con las patas posteriores, resultando a veces en una reduccion del tiempo de copula y de transferencia de espermatozoides.
Las hembras de S. cincticeps pertenecientes al tratamento T1 o T2 presentaron semejanza en el porcentaje de huevos viables indicando que, aparentemente, no hubo una reduccion en la cantidad de espermatozoides transferidos. Esto concuerda con lo encontrado para P. nigrispinus (Torres et al. 1997) y puede ser un atributo adecuado para que esas chinches se establezcan en un ecosistema agricola o forestal. Sin embargo, en arañas, la hembra de S. lineatus confinada al macho tuvo efecto deletereo en la produccion total de descendientes (Maklakov et al. 2005). Estos resultados sugieren que organismos poliandricos presentan diferentes comportamientos cuando la pareja se interrelaciona y que dos o mas apareamientos no necesariamente implican un aumento en la eficacia reproductiva de algunas hembras. Ademas, otros aspectos biologicos pueden ser allerados en las hembras debido a la presencia constante del macho pudiendo reducir la actividad de alimentacion como se observo en Lampetis nigerrima (Kerremans) (Coleoptera: Buprestidae) (De Nadai 2005) y consecuentemente disminuir el exito reproductivo (Parra 1991). De acuerdo con Arnqvist y Nilsson (2000) el efecto de multiples copulas todavia no es bien entendido y es dificil de predecir, pues los diferentes componentes reproductivos de las hembras son afectados de forma negativa y/o positiva por varios factores como el propio aparcamiento, la transferencia adecuada de esperma y las sustancias presentes en el eyaculado.
Al analizarse el comportamiento de eclosion por puesta, se observo que hubo mayor numero de huevos infertiles en las ultimas puestas para las hembras del tratamiento T2, lo que parece ser semejante entre especies poliandricas como las chinches predadoras (Torres et al. 1997; Torres y Zanuncio 2001). Este resultado indica que, probablemente, las hembras de S. cincticeps no lograron obtener y almacenar en la espermateca la cantidad de espermatozoides suficientes para fertilizar todos sus ovulos. Este resultado no se encontro en las hembras del tratamiento T1 (confinadas junto al macho durante todo su periodo de reproduccion), las cuales mantuvieron una alta fertilidad a lo largo de su periodo reproductivo, demostrando claramente el efecto positivo de multiples apareamientos en la fertilidad de las hembras (Arnqvist y Nilsson 2000). Por lo tanto, la transferencia efectiva de espermatozoides debe evaluarse comparandose la eclosion de los huevos en funcion del orden de puesta y no compararse apenas el promedio de huevos viables debido a que eso puede enmascarar los resultados. Ademas de ese tipo de beneficio, es posible que las hembras tambien reciban junto con el eyaculado otras sustancias como agua y nutrientes (Arnqvist et al. 2005).
Si bien en grupos relacionados, como algunas especies del genero Podisus, se ha registrado canibalismo (Richman y Whitcomb 1978), esto no se observo en S. cincticeps. La presencia del macho durante toda la vida de las hembras de S. cincticeps no tuvo ese efecto negativo debido a que estas presentaron promedio de longevidad semejante (alrededor de 26 dias) en todos los tratamientos. Estos resultados son proximos a aquellos encontrados por Zanuncio et al. (1992) quienes observaron una longevidad de 30 dias para hembras de esa misma especie a 25ºC en condiciones de laboratorio.
En conclusion, hembras y machos de S. cincticeps pueden mantenerse en una proporcion 1:1 en las colonias de laboratorio, por lo menos hasta alcanzar 25 dias de edad, debido a que durante ese periodo las chinches acumularon, aproximadamente, 80% de la productividad de huevos. De esa forma se optimizaria la cria de ese predador en laboratorio y consecuentemente el establecimiento de programas de control biologico utilizando ese tipo de enemigo natural. Ademas el hecho de confinar las parejas de S. cincticeps no influyo negativamente en el desempeño reproductivo de las hembras y contribuyo para mantener una alta viabilidad a lo largo de su periodo reproductivo.
Agradecimientos
Los autores agradecen al Consejo Nacional de Desarrollo Cientifico y Tecnologico (CNPq), a la Fundacion de Amparo a la Investigacion del Estado de Minas Gerais (FAPEMIG) y a la Federacion de Industrias del Estado de Minas Gerais (FIEMG) por el apoyo Financiero recibido para la realizacion de este experimento.
Literatura citada
ARNQVIST, G; NILSSON, T. 2000. The evolution of polyandry: multiple mating and females fitness in insects. Animal Behaviour 60 (2): 145-267. [ Links ]
ARNQVIST G.; NILSSON, T; KATVALA, ML 2005. Mating rale and fitness in female bean weevils. Behavioral Ecology 16(1): 123-127. [ Links ]
DE CLERCQ, P.; DEGHEELE, D. 1997. Effects of mating status on body weight. imposition, egg load, and predation of the predatory stinkbug Podisus maculiventris (Hemiplera: Pentatomidae). Annals of the Entomological Society of America 90 (2): 12 N127. [ Links ]
DE CLERCQ. P.; WYCKHUYS. K.: OLL-VEIRA, H.N.; KLAPWIJK, J. 2002. Predation by Podisus maculiveniris on different life stages of Nezara vii idula. Florida Entomologist 85 (1): 197-202. [ Links ]
DE NADAI. J. 2005. Biologia de Lampetis nigerrima (Kerremans, 1897) (Coleoplera: Buprestidae) em eucalipto. Tesis de Maestria en Entomologia. Universidade Federal de Viçosa, Viçosa. 53 p. [ Links ]
GOMES, F.P. 1978. Curso de estatistica experimental. ESALQ-USP, Piracicaba. Sao Paulo. Brasil.430p. [ Links ]
MAKLAKOV. A. A.: BILDE. T.; LUBIN. Y. 2005. Sexual conflict in the wild: elevated mating rates reduces female lifetime reproductive success. American Naturalist 165(5): S38-S45. [ Links ]
MOLINA-RUGAMA. A.J.; ZANUNCIO, J.C.: TORRES. J.B.: ZANUNCIO. T.V. 1997. Longevidad y fecundidad de Podisus nigrispinus (Heteroptera: Pentatomidae) alimentado con Muse a domesticu L. (Diptera: Muscidac) y frijol. Revista de Biologia Tropical 45 (3): 1125-1130. [ Links ]
MOLINA-RUGAMA, A.J.; ZANUNCIO, J.C.: ZANUNCIO. TV.: DE OLIVEIRA, M.L.R. 1998. Reproductive strategy of Podisus rostral is (Stal) (Heteroptera: Pentatomidae) females under different feeding intervals. Biocontrol Science and Technology 8 (4): 583-588. [ Links ]
MOURAO, S.A.; J.C. ZANUNCIO; A.J. MOLINA-RUGAMA; E.F. VILELA; M.C. LACERDA. 2003. Efeitodaescassez de presas na sobrevivencia e rcproducao do predador Supputius cincticeps (Stal) (Heteroptera: Pentatomidae). Neotropical Entomology 32 (2): 469-473. [ Links ]
PARRA. J.R.P. 1991. Consumo e utilizacao de alimentos por insetos. p. 9-66. En: Panizzi, A.R.; Parra, J.R.P. (eds.). Ecologia nutricional de insetos e suas implicates no manejo de pragas. Ed. Manole. Sao Paulo. Brasil. 359 p. [ Links ]
RICHMAN, D.B.; WHITCOMB. W.H. 1978. Comparative life cycles of four species of predatory stinkbugs (Hemiplera: Pentatomidae). Florida Entomologist 61 (3): 113-119. [ Links ]
RODRIGUEZ. R.L.S. 1998. Possible female choice during copulation in Ozophora baranowskii (Heteroptera: Lygaeidae): female behavior, multiple copulations, and sperm transfer. Journal of Insect Behavior 11 (5): 725-741. [ Links ]
TORRES, J.B.; ZANUNCIO, J.C; OLIVEIRA. M.C. 1997. Mating frequency and its effect on female reproductive output in the stinkbug predator Podisus nigrispinus (Heteroptera: Pentatomidae). Medede-lingen Fueulteit Landbouwwetenschappen Universsteit Gent. 62 (2b): 491 -498. [ Links ]
TORRES. J.B.; ZANUNCIO. J.C. 2001. Effects of sequential mating by males on reproductive output of the stinkbug predator Podisus nigrispinus. BioControl 46 (4): 469-480. [ Links ]
ZANUNCIO, T.V.; ZANUNCIO. J.C: VILELA, E.F. 1992. Aspectos biologicos da fase adulta de Supputius cincticeps Stal. 1860 (Hemiplera: Pentatomidae). predador de lagartas desfolhadoras de eucalipto. Instituto de Pesquisas e Estudos Floreslais 45 (1): 35-39. [ Links ]
ZANUNCIO, J.C; ALVES. J.B.; ZANUNCIO, TV: GARCIA. J.F. 1994. Hemipterous predators of eucalypt defoliator caterpillars. Forest Ecology and Management 65 (I): 65-73. [ Links ]
ZANUNCIO. J.C; SAAVEDRA, J.L.D.; OLIVEIRA, H.N.; DEGHEELE, D.; DE CLERCQ. P. 1996. Development of the predatory stinkbug Brontocoris tabidus (Signoret) (Heteroptera: Pentatomidae) on different proportions of an artificial diet and pupae of Tenebrio molitor L. (Coleoptera: Tenebrionidae). Biocontrol Science and Technology 6 (4): 619-625. [ Links ]
ZANUNCIO, J.C; SAAVEDRA. J.L.D.; ZANUNCIO, T.V; SANTOS, G.P. 1996-1997. Desarrollo y reproduction de Supputius cincticeps (I ieteroplera: Pentalomidae) en diela artificial por dos generaciones. Revista de Biologia Tropical 44 (3) /45 (1): 247-251. [ Links ]
ZANUNCIO. J.C; MOLINA-RUGAMA, A.J.; SERRAO. J.E.: PRATISSOLI. D. 2001. Nymphal development and repro duction of Podisus nigrispinus (Heterop tera: Pentatomidae) fed with combinations of Tenebrio molitor (Coleoplera: Tenebrio nidae) pupae and Musca domesticu (Diptera: Muscidac) larvae. Biocontrol Science and Technology 1 1 (3): 331-337. [ Links ]
ZANUNCIO. J.C; MOLINA-RUGAMA, A.J.; SANTOS. G.P.; RAMALHO. F.S. 2002. Effect of body weight on fecundity and longevity of the stinkbug predator Podisus rostralis. Pesquisa Agropecuaria Brasileira 379): 1225-1230. [ Links ]
Recibido:01-sep-04 - Aceptado: 16-jul-05

