










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488
Rev. Colomb. Entomol. vol.39 no.1 Bogotá Jan./June 2013
Preferencia de oviposición del barrenador del amaranto, Aerenea quadriplagiata (Coleoptera, Cerambycidae) en condiciones de campo
Oviposition preference of the amaranth stem borer, Aerenea quadriplagiata (Coleoptera, Cerambycidae) under field conditions
María Begoña Riquelme V.1,2, María Agustina Ansa1,3 y Marina Vilma Santadino1,4
1 Departamento de Tecnología, Universidad Nacional de Luján. Rutas Nacionales 5 y 7, (6700) Luján, Buenos Aires, Argentina.
2 Doctora en Cs. Biológicas. Laboratorio de Zoología Agrícola. mbriquelme@hotmail.com. Autor para correspondencia.
3 Ingeniera agrónoma. Laboratorio de Conservación del Sistema Agropecuario. agus3287@hotmail.com.
4 Ingeniera agrónoma. Laboratorio de Zoología Agrícola. msantadino@yahoo.com.ar.
Recibido: 21-nov-2012 - Aceptado: 23-may-2013
Resumen: El amaranto es considerado como un "cultivo estratégico" debido a su variabilidad genética, su rusticidad y su valor nutritivo. Una de sus principales adversidades fitosanitarias en la Región Pampeana argentina es el cerambícido barrenador, Aerenea quadriplagiata. Este trabajo tuvo como objetivo evaluar la preferencia de oviposición de este insecto sobre distintos cultivares y características morfométricas de la planta amaranto. En el campo experimental de la Universidad Nacional de Luján, se sembraron durante dos años consecutivos, parcelas con diferentes cultivares de Amaranthus. Se relacionaron la presencia y el número de oviposiciones y la proporción de oviposiciones efectivas (con desarrollo de larvas) con el cultivar, la altura y el diámetro de las plantas. El cultivar hortícola presentó la menor proporción de oviposiciones efectivas, el menor porcentaje de plantas con oviposiciones y el mayor diámetro con un 50% de probabilidades de ser ovipuesto. Estos resultados sugieren que la preferencia de las hembras del barrenador está influenciada tanto por el cultivar como por el diámetro de la planta y que A. mantegazzianus L. cv hortícola es la variedad más aconsejada en zonas con una alta incidencia del barrenador.
Palabras clave: Amaranthus. Interacción planta-insecto. Insecto endofitófago.
Abstract: Amaranth is currently considered a "strategic grain crop" because of its broad genetic variability, hardiness, and nutritional value. In the Argentinean Pampas Region, one of its major pests is the cerambycid stemborer, Aerenea quadriplagiata. This study aimed to evaluate the oviposition preference of this pest related to both different cultivars and morphometric characteristics of the amaranth plant. In the experimental field of the Universidad Nacional de Luján, plots with different cultivars of Amaranthus were planted in two consecutive years. The presence and number of oviposition and oviposition effective ratio (with development of larvae) were related to the cultivar, the height and the stem diameter. The horticultural cultivar presented the lowest oviposition effective ratio, the lowest percentage of plants with oviposition and larger stem diameter with a 50% chance of being oviposited by A. quadriplagiata. These results suggest that the behavior of female stem borer is influenced by both the cultivar and stem diameter and that A. mantegazzianus L. cv horticola is the most recommended cultivar in areas where there is high incidence of the stem borer.
Key words: Amaranthus. Insect-plant interaction. Endophytophagous insect.
Introducción
El género Amaranthus L. abarca más de setenta especies vegetales, la mayoría de las cuales son nativas de América y se cultivan desde mucho antes de la colonización española. Históricamente en Argentina, fuera del cultivo practicado por los pueblos originarios, los amarantos graníferos solo habían sido cultivados desde principios del siglo pasado en parcelas experimentales o en pequeños lotes asociados con maíz en las provincias de Jujuy, Salta, Tucumán y Catamarca (Covas 1994). Sin embargo, este panorama ha ido cambiando debido a su promoción como cultivo alternativo para la Región Pampeana semiárida, el Noroeste Argentino y la Región Patagonia Norte, por su elevada adaptabilidad a diversas condiciones ambientales y el alto valor nutritivo de sus hojas y granos (Covas 1994; Martín de Troiani et al. 2005).
Entre las adversidades fitosanitarias asociadas a este cultivo, se han documentado varios insectos con hábito barrenador (Vilca y Jeri 1991; Phogat et al. 1994; Louw et al. 1995; Banjo 2007; Tara et al. 2009; Pérez Torres et al. 2011). Dado su régimen alimentario, los barrenadores ingresan al tallo y realizan galerías en el mismo, dificultando la circulación de agua y nutrientes (Arguedas 2006). Aunque en general se ha observado que los daños asociados a estos insectos permiten que las plantas de amaranto se desarrollen por completo (Ansa, 2012) y no disminuya el rendimiento del cultivo (Torres Saldaña et al. 2004), en otros casos se produce una elevada proporción de panojas y plantas quebradas que podrían afectar la producción (Martínez y Riquelme Virgala 2007).
En Argentina se han citado dos coleópteros como principales plagas de hábito barrenador para el cultivo de amaranto. En la provincia de La Pampa, una especie del género Conotrachelus (Schöngerr, 1845) (Curculionidae) presenta una gran incidencia (Niveyro et al. 2008), mientras que en la provincia de Buenos Aires, un barrenador de la familia Cerambycidae Aerenea quadriplagiata (Boheman, 1859), fue registrado en varios estudios en Luján (De Haro y Martínez 1995; Sbravatti 1999; Cordo et al. 2004) y en otras localidades de la misma provincia y de la provincia de Mendoza (Di Iorio 2003, 2006). Posteriormente, Martínez y Riquelme Virgala (2007) confirmaron que A. quadriplagiata es la principal plaga del cultivo de amaranto en el noreste bonaerense por su elevada incidencia y sus daños.
Diversos factores inciden en la selección de la planta por un insecto herbívoro (Bernays 1994). Para los insectos de hábitos endofitófagos como los cerambícidos, son las hembras adultas las que a través de señales físicas y químicas deben encontrar y seleccionar la planta adecuada para oviponer, en cuanto que las larvas no tendrán la capacidad de dispersarse en busca de nuevos hospedantes. Estas señales pueden estar dadas por el material genético y las características morfológicas de la planta (Paulino Neto 2005; Marquina et al. 2006), factores que podrían ser manejados a través de la elección de la especie y del cultivar y la distribución espacial del cultivo (Niveyro y Salvo 2012). Este trabajo tuvo como objetivo evaluar la preferencia de oviposición del barrenador del amaranto asociado a distintos cultivares y características morfométricas de la planta, con el fin de buscar herramientas para el manejo integrado de esta plaga.
Materiales y métodos
Las observaciones se realizaron en parcelas de amaranto ubicadas en el campo experimental de la Universidad Nacional de Luján, provincia de Buenos Aires (latitud 34º36"S y longitud 59º04"O, a 28 msnm), en las temporadas 2010/11 y 2011/12. Las especies y cultivares empleados fueron: Amaranthus mantegazzianus L. cv. hortícola, A. hypochondriacus L. cv. artasa 9122, A. cruentus L. cv. don guiem, A. cruentus L. cv. mejicano. Las observaciones se realizaron cuando los cultivares se encontraban en la etapa fenológica de fructificación. El diseño experimental de ambos ensayos fue en bloques completos aleatorizados con tres repeticiones y cada parcela constó de tres (primer ciclo) y cinco (segundo ciclo) líneas de 3 m de largo, con una distancia entre surcos de 70 cm. Las parcelas se sembraron a chorrillo, el control de malezas fue manual y no se realizaron raleos ni tratamientos fitosanitarios.
Primer ciclo de cultivo. Se tomaron muestras de diez plantas con síntomas de oviposición por parcela. El daño por oviposición en esta especie es muy característico, la hembra realiza un pequeño corte transversal para colocar un único huevo en la parte apical del orificio y luego el tejido que se encuentra alrededor de la herida se torna castaño oscuro (Ansa 2012).
En laboratorio, a cada planta se le midió el diámetro a los 20 cm del cuello, la altura hasta la base de la panoja y se contabilizó el número de oviposiciones. El diámetro del tallo se determinó empleando un calibre automático de 0,01 mm de precisión y, posteriormente, se diseccionó cada tallo con un bisturí para observar la presencia de galerías.
Se registraron las siguientes variables: N° de oviposiciones por planta (O), N° de galerías de más de 1 cm por planta (G) y porcentaje de oviposiciones efectivas (OE) definido como: OE (%) = (G/O)*100. Cabe aclarar que el porcentaje de oviposiciones efectivas representa los casos en los que la larva se desarrolló y, por lo tanto, se evidenció el daño, sin diferenciar si la mortalidad ocurrió en el estado de huevo o de larva neonata. Se realizó un análisis de varianza para comprar cada una de estas variables entre los cultivares (tratamiento) y en los casos en los que resultó significativo, se separaron las medias a posteriori con la prueba de Tuckey. Para evaluar si el número de oviposiciones por planta se relaciona linealmente con las características morfológicas de la misma se realizaron, para cada cultivar por separado, los análisis de correlación correspondientes para relacionar a cada variable (O, G y OE) con el diámetro y la altura. Para ello se estimó el coeficiente de correlación lineal de Pearson y su significancia (Zar 1996).
Segundo ciclo de cultivo. Para evaluar la preferencia de oviposición entre cultivares, se eligieron al azar diez plantas por parcela en las que se registró el número de plantas con al menos una oviposición y el diámetro del tallo a 20 cm del cuello. Se realizó un ANCOVA con la variable porcentaje de plantas con oviposición, considerando como tratamiento a los cultivares y como covariable al diámetro medio de las plantas de la muestra y se separaron las medias con la prueba de Tuckey. Posteriormente, se hizo un análisis de regresión logística para cada cultivar con la variable binomial: con oviposición vs. sin oviposición, cuya variable independiente fue el diámetro de la planta. La significancia se estudió utilizando el estadístico de Wald. Posteriormente se calculó el odds yel diámetro de la planta con un 50% de chances de ser oviposiuesto. El odds es un parámetro quecorresponde a las chances que tiene de ocurrir el evento estudiado por cada incremento de la variable independiente (Agresti 1996).
En todos los análisis estadísticos se tuvo en cuenta un error estadístico del 5% y se utilizaron los módulos ANOVA/MANOVA, MULTIPLE REGRESSION y GZM (Generalized Linear Models) del programa estadístico STATISTICA (versión 2000).
Resultados y discusión
Efecto de los cultivares. El número de oviposiciones y el de galerías por planta (Fig. 1) no presentaron diferencias significativas entre los cultivares (Oviposición: F3,6= 3,91; P = 0,07; Galerías: F3,6= 0,12; P = 0,94). Sin embargo, cuando ambas variables se relacionaron para obtener el porcentaje de oviposiciones efectivas, este índice sí reflejó diferencias estadísticas (F3,6= 3,23; P = 0,005). El cultivar hortícola presentó el menor valor, con solo un 30% de oviposiciones en las que prosperó el desarrollo de la larva (Fig. 1). Esto podría estar asociado a una menor calidad nutricional del tejido o a la presencia de metabolitos secundarios que limiten el desarrollo inicial del barrenador.
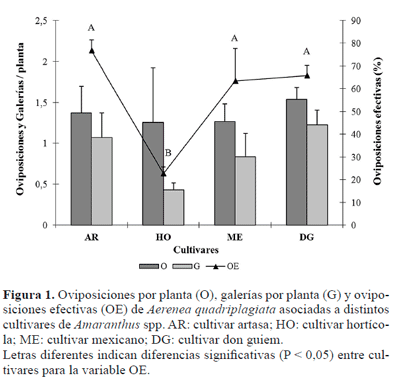
Existen antecedentes, para otras especies de cerambícidos xilófagos, de que las plantas pueden defenderse de su ataque produciendo sustancias resinosas o de origen polifenólico que no permiten la emergencia del huevo y/o el desarrollo de la larva (Brugnoni 1980) o que otras condiciones como el contenido de humedad de los tejidos pueden ser fundamentales para la supervivencia de las mismas (Hanks et al. 1999).
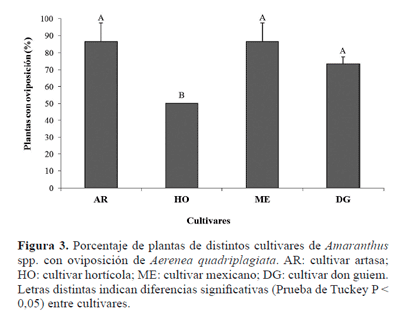
En el segundo ensayo el % de plantas con oviposición fue estadísticamente diferente entre los materiales genéticos (F3,7= 5,87; P = 0,025). Del mismo modo, Torres Saldaña et al. (2004) observaron en México que el número de larvas y el porcentaje de barrenación por Hypolixus truncatulus (F.) (Coleoptera, Curculionidae) y Amauromyza abnormalis (Malloch, 1913) (Diptera, Agromyzidae), fueron significativamente diferentes entre los dos cultivares de amaranto que estudiaron. El cultivar hortícola fue el menos preferido por la hembra del barrenador, mostrando diferencias significativas con los demás cultivares (Prueba de Tuckey P < 0,05, Fig. 3). Este resultado junto con el menor porcentaje de oviposiciones efectivas observado para este cultivar en el ciclo anterior, sugieren una relación entre la selección de las hembras al oviponer y la supervivencia inicial de su descendencia. Este comportamiento ha sido observado en otras especies de cerambícidos (Hanks et al. 1995; Morewood et al. 2003) y concuerda con una de las hipótesis ecológicas que relaciona la preferencia de oviposición con el mejor desempeño de la progenie (Thompson 1987). Además, se ha visto que la probabilidad de que esta hipótesis se cumpla es mayor en aquellos insectos con oviposiciones endofíticas, debido a que insertar los huevos en el tejido vegetal en el que las larvas se desarrollarán, ofrece la oportunidad de detectar las señales de las plantas que indican la calidad del sitio de oviposición (Price 1994).
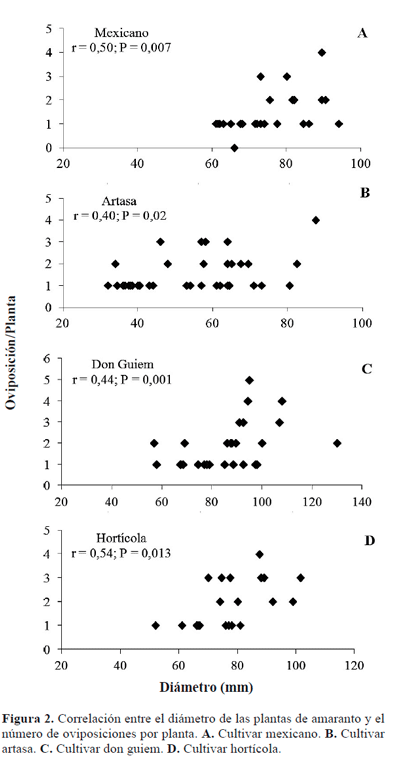
Efecto de las características morfométricas de la planta. La altura de las plantas no se relacionó con ninguna de las variables, lo que sugiere que esta característica morfológica no influye sobre el ataque del barrenador, mientras que la correlación fue positiva y estadísticamente significativa para la relación entre el diámetro con el número de oviposiciones por planta en todos los cultivares (Fig. 2). En concordancia, durante el segundo ensayo la presencia de oviposiciones en la planta estuvo significativamente afectada por el diámetro en todos los cultivares. El coeficiente de regresión logística fue positivo en todos los casos (Tabla 1), lo que significa que a medida que aumenta el diámetro del tallo, aumentan las probabilidades de que una planta sea ovipuesta por el barrenador. De esta manera, el diámetro parece jugar un papel de relevancia sobre la selección de la hembra al oviponer y sobre el número de huevos que ésta coloca en una misma planta, lo que ha sido observado en otras especies de cerambícidos (Shibata et al. 1994; Diodato et al. 1997; Suárez Hernández 2003; Paulino Neto et al. 2005). Específicamente en el cultivo de amaranto, Niveyro y Salvo (2012) observaron una menor incidencia de gorgojos barrenadores en A. hypochondriacus cv artasa cuando fue cultivado a alta densidad, lo que estos autores relacionaron al menor diámetro de los tallos con respecto a cultivos sembrados menos densos. Sin embargo, estos resultados se contradicen con los observados por Torres Saldaña et al. (2004), quienes encontraron que el diámetro de las plantas de amaranto no influyó sobre la incidencia de los barrenadores H. truncatulus y A. abnormalis.
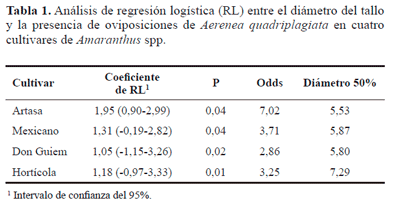
El diámetro del tallo con un 50% de probabilidad de presentar oviposición varió entre 5,53 y 7,29 mm para artasa y horticola, respectivamente. De manera similar, Sbravatti (1999) observó que el barrenado de tallos por A. quadryplagiata se presenta a partir de los 6 mm de diámetro del tallo de amaranto. El diámetro del tallo depende en gran medida del material genético y de la densidad de siembra, por lo que conocer el diámetro susceptible para cada cultivar es un dato relevante para el manejo de la distribución espacial del cultivo.
Conclusiones
Los resultados de este trabajo indicarían que las hembras de Aerenea quadriplagiata prefieren las plantas para oviponer tanto por características propias del material genético como por el diámetro del tallo y que esta selección podría estar relacionada a la mayor supervivencia de su progenie en dichos materiales. Estas observaciones se podrían resumir en el comportamiento del barrenador asociado al cultivar Hortícola. Este cultivar fue en principio el material genético menos elegido para oviponer, luego las hembras sólo lo seleccionan si el diámetro de tallo es aproximadamente un 30% mayor que el de los demás cultivares y por último la supervivencia de la progenie se ve significativamente reducida. Por lo tanto, el material A. mantegazzianus cv. hortícola es la variedad más aconsejada en zonas en donde existe una alta incidencia del barrenador. Futuros estudios relacionados a la fecha y densidad de siembra serían recomendables para reducir en mayor medida la incidencia del barrenador.
Agradecimientos
Agradecemos especialmente a la Ingeniera Carmen Martínez por iniciarnos en este tema en el que ella fue pionera. Agradecemos además al equipo docente de Conservación del Sistema Agropecuario por la provisión de las semillas, obtenidas en el marco del proyecto de extensión universitaria "Proyecto de promoción, desarrollo e integración asociativa de productores vinculados a producciones alternativas en la Cuenca del Río Luján y sus alrededores" y por las tareas de siembra y mantenimiento de las parcelas.
Literatura citada
AGRESTI, A. 1996. An introduction to categorical data analysis. Wiley & Sons, Inc. University of Florida, EEUU. 293 p. [ Links ]
ANSA, A. 2012. Observaciones sobre la entomofauna asociada a cinco cultivares de amaranto (Amaranthus spp.) cultivados. Tesis (ingeniero agrónomo). Universidad Nacional de Luján, Argentina. 47 p. [ Links ]
ARGUEDAS, M. 2006. Clasificación de tipos de daños producidos por insectos forestales. Segunda parte. Revista Forestal Costa Rica 3 (9): 1-8. [ Links ]
BANJO, A. D. 2007. Bio-ecology and life history of Gasteroclisus rhomboidalis (Coleoptera: Curculionidae) a pest of Amaranthus cruentus (L.) Sauer. Journal of Entomology 4 (4): 308-316. [ Links ]
BERNAYS, E.; CHAPMAN, R. 1994. Host-plant selection by phytophagous insects. Chapman & Hall, New York. 296 p. [ Links ]
BRUGNONI, H. 1980. Plagas forestales. Editorial Hemisferio Sur, Buenos Aires, Argentina. 216 p. [ Links ]
CORDO, H. A.; LOGARZO, G.; BRAUN, K.; DI IORIO, O. 2004. Catálogo de insectos fitófagos de la Argentina y sus plantas asociadas. Sociedad Entomológica Argentina Ediciones. Buenos Aires, Argentina. 734 p. [ Links ]
COVAS, G. 1994. Perspectivas del cultivo de los amarantos en la República Argentina. EE INTA Anguil, La Pampa, Argentina. Publicación miscelánea Nº13. 10 p. [ Links ]
DE HARO, A. M.; MARTÍNEZ, C. L. 1995. Especies animales asociadas al cultivo de Amaranthus sp. En: Actas del III Congreso Argentino de Entomología, Mendoza, Argentina, p. 165. [ Links ]
DI IORIO, O. R.; RIQUELME, A. H.; ESTEBAN-DURAN, J. R. 2003. Cerambycidae (Coleoptera) of the province of Mendoza, Argentina. Spanish Journal of Agricultural Research 1 (4): 15-29. [ Links ]
DI IORIO, O. R.; FARINA, J. 2006. La fauna de Cerambycidae (Coleoptera) de la provincia de Buenos Aires, Argentina. Revista del Museo Argentino de Ciencias Naturales 8 (2): 261-287. [ Links ]
DIODATO, L; DARCHUK, E.; NOTARIO, A.; CASTRESANA, L. 1997. Estudio sobre el comportamiento de oviposición del "cortapalos" Oncideres guttulata Thompson (Coleoptera, Cerambycidae) sobre "quebracho colorado", Schinopsis quebracho colorado (Shelecht) Bark et Meyer. Boletín Sanidad Vegetal Plagas 23: 257-261. [ Links ]
HANKS, L. M.; MILLAR, J. G.; PAINE, T. D. 1995. Biological constraint on host-range expansion by the wood-boring beetle Phoracantha semipunctata (Coleoptera: Cerambycidae). Annals of the Entomological Society of America 88 (2): 183-188. [ Links ]
HANKS, L. M.; PAINE, T. D.; MILLAR, J. G.; CAMBELL, CH. D.; SCHUCH, U. K. 1999. Water relations of host trees and resistance to the phloem-boring beetle Phoracantha semipunctata F. (Coleoptera: Cerambycidae). Oecologia 119: 400-407. [ Links ]
LOUW, S.; VAN EEDEN, C. F.; WEEKS, W. J. 1995. Curculionidae (Coleoptera) associated with wild and cultivated Amaranthus spp. (Amarantaceae) in South Africa. African Crop Science Journal 3 (1): 93-98. [ Links ]
MARTÍN de TROIANI, R.; REINAUDI, N.; TASSONE, M. 2005. El amaranto en su mesa. En: Actas del X Cong. Arg. de Ciencia y Tecnología de Alimentos, Mar del Plata, Argentina. Disponible en: http://www.agro.unlpam.edu.ar/publicaciones-pdf/El%20amaranto%20en%20su%20mesa.pdf [Fecha revisión: 5 agosto 2012] [ Links ].
MARTÍNEZ, C. L.; RIQUELME VIRGALA, M. B. 2007. Fauna asociada a algunos cultivos hortícolas, aromáticos, medicinales y condimentarios. Horticultura Argentina 26 (61): 101. [ Links ]
MOREWOOD, W. D.; NEINER, P. R.; McNEIL, J. R.; SELLMER, J. C.; HOOVER, K. 2003. Oviposition preference and larval performance of Anoplophora glabripennis (Coleoptera, Cerambycidae) in four eastern North American hardwood tree species. Environmental Entomology 32 (5): 1028-1034. [ Links ]
NIVEYRO, S.; SALVO, A. 2012. Prácticas culturales exitosas para disminuir el ataque de insectos barrenadores de Amaranthus hypocondriacus. Libro de Resúmenes del VIII Congreso Argentino de Entomología, p. 288. [ Links ]
NIVEYRO, S.; BAUDINO, E.; FALKENSTEIN, L.; SÁENZ, C. 2008. Assessment of the arthropods associated with the amaranth crop in the central Argentina. Proceedings V Internacional Symposium of the European Amaranth Association, Slovak Republic, p. 44-47. [ Links ]
PAULINO NETO, H. F.; ROMERO, G.; VASCONCELLOS NETO, J. 2005. Interactions between Oncideres humeralis Thompson (Coleoptera, Cerambycidae) and Melastomataceae: Hos-plant selection and patterns of host use in Southeast Brazil. Neotropical Entomology 34 (1): 7-14. [ Links ]
PÉREZ TORRES, B. C.; ARAGÓN GARCÍA, A.; PÉREZ AVILÉS, R.; HERNÁNDEZ, L. R.; LÓPEZ OLGUÍN, J. F. 2011. Estudio entomofaunístico del cultivo de amaranto (Amaranthus hypocondriacus L.) en Puebla, México. Revista Mexicana de Ciencias Agrícolas 2 (3): 359-371. [ Links ]
PHOGAT, B. S.; BHALLA, S.; MAL, B. 1994. Seasonal incidence of stem weevil (Hypolyxus truncatulus) and its effect on growth and grain yield of amaranth (Amaranthus hypocondriacus). Indian Journal of Agricultural Sciences 64: 261-262. [ Links ]
PRICE, P. W. 1994. Phylogenetic constraints, adaptive syndromes, and emergent properties: from individuals to population dynamics. Researches on Populations Ecology 36: 1-12. [ Links ]
SBRAVATTI, A. M. 1999. Aspectos biológicos de Aerenea quadriplagiata (Boheman 1858) (Coleoptera, Cerambycidae) barrenador del tallo de Amaranthus spp. Tesis (ingeniero agrónomo). Universidad Nacional de Luján, Argentina. 75 p. [ Links ]
SHIBATA, E.; WAGUCHI, Y.; YONEDA, Y. 1994. Role of tree diameter on the damage caused by the sugi bark borer (Coleoptera, Cerambycidae) to the Japanese cedar, Cryptomeria japonica. Environmental Entomology 23 (1): 76-79. [ Links ]
SUÁREZ HERNÁNDEZ, L. A. 2003. Prospección del daño ocasionado por dos insectos xilófagos asociados a renovales de Notophagus en las provincias de Valdivia, Osorno y Llanquihue, Décima Región. Tesis (ingeniero forestal). Universidad Austral de Chile. 47 p. [ Links ]
TARA, J. S.; AZAM, M.; AYRI, S.; FEROZ, M.; RAMAMURTHY, V. 2009. Bionomics of Hypolixus truncatulus (Coleoptera, Curculionidae, Lixinae, Lixini), a major pest of Amaranthus caudatus L. Munis Entomology & Zoology 4 (2): 510-518. [ Links ]
THOMPSON, J. 1987. Evolutionary ecology of the relationship preference and performance of offspring in phytophagous insects. Entomolgia Experimentalis et Applicata 47: 3-14. [ Links ]
THOMPSON, J.; PELLMYR, O. 1990. Evolution of oviposition behavior and host preference in Lepidoptera. Annual Review of Entomology 36: 65-89. [ Links ]
TORRES SALDAÑA, G.; TRINIDAD SANTOS, A.; TRUJILLO, T. R.; CASTILLO JUÁREZ, H.; BAUTISTA MARTINEZ, N.; DE LEON GONZALEZ, F. 2004. Barrenación del tallo de amaranto por Hypolixus trucatulus (Coleoptera: Curculionidae) y Amauromyza abnormalis (Diptera: Agromyzidae). Acta Zoológica Mexicana 20 (1): 131-140. [ Links ]
VILCA, J.; JERI, A. 1991. Daños del barrenador del tallo (Coleoptera: Curculionidae) en germoplasma de "Achita" (Amaranthus caudatus L.) en Ayacucho. En: Actas del VII Congreso Internacional sobre cultivos andinos, La Paz, Bolivia, 1991, pp. 159-161. [ Links ]
ZAR, J. H. 1996. Biostatistical analysis. Prentice Hall, New Jersey, EEUU. 918 p. [ Links ]
