










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.20 no.1 Bogotá Jan./Apr. 2015
https://doi.org/10.15446/abc.v20n1.42260
Artículo de revisión
USE OF FOOD RESOURCES BY DETRITIVOROUS FISH IN FLOODPLAINS: A SYNTHESIS
El uso de los recursos alimenticios por peces detritívoros en planicies de inundación: una síntesis
Ana Rute AMADEU SANTANA1, Martin WERTH2, Evanilde BENEDITO-CECILIO3.
1Departamento de Biología. Programa de Pós-graduação em Biologia Comparada. Universidade Estadual de Maringá (UEM), Brasil.
2Institute of Systematic Botany and Ecology. Ulm University, Germany.
3Núcleo de Pesquisas em Limnologia, Ictiologia e Aqüicultura (Nupelia). Universidade Estadual de Maringá (UEM), Brasil.
For correspondence: eva@nupelia.uem.br
Received 24th February 2014, Returned for revision 29th April 2014, Accepted 5th June 2014.
Citation/Citar este artículo como: Amadeu Santana AR, Werth M, Benedito-Cecilio E. Use of food resources by detritivorous fish in floodplains: a synthesis. Acta biol. Colomb. 2015;20(1):5-14. doi: http://dx.doi.org/10.15446/abc.v20n1.42260
ABSTRACT
The present review investigated the sources of energy for detritivorous fish species in floodplains. Recent studies have shown that environmental changes that alter the terrestrial vegetation affect essentially the resource base in these areas, such as the detritus. In aquatic environments, especially floodplains, a major part of metabolic processes is supported by terrestrial detritus, which accounts for maintaining the detritivorous food chain. Thus, one way to evaluate environments changes is through isotopic studies of organic matter, which allows acquiring information about the quantity and quality of organic matter. This approach is required for understanding the interactions between the organic matter dynamics and its ecological effects, allowing the monitoring of these resources. However, researches on the energy flow in food chains should also employ an approach that quantifies the magnitude of these food subsidies, as also the access routes of aquatic consumers, in watersheds of different sizes and in diverse land uses.
Keywords: environmental changes, detritus, microbial biomass, stable isotopes.
RESUMEN
Esta revisión procuró investigar las fuentes de energía para las especies de peces detritívoras en la llanura de inundación. Los estudios han demostrado que las transformaciones ambientales que causan los cambios en la vegetación terrestre afectan principalmente la base de los recursos en estos planos inundables, tales como el detritus. En los ambientes acuáticos, especialmente las llanuras de inundación, la mayoría de los procesos metabólicos están sustentados por el detritus terrestre, y esta función es responsable de mantener las redes detritívoras. Una forma de evaluar estos cambios es a través de estudios isotópicos de la materia orgánica, los cuales permiten obtener información sobre su cantidad y su calidad mediante el análisis de la biomasa microbiana. Este enfoque es necesario para comprender las interacciones entre la dinámica de la materia orgánica y sus efectos ecológicos, lo que permite a su vez la valoración de estos recursos. Sin embargo, se concluye que los estudios futuros también deben cuantificar la magnitud de estos subsidios energéticos, asícomo las vías de acceso de los consumidores acuáticos, lo cual debe evaluarse en cuencas hidrográficas de diferentes tamaños y con diversos usos de la tierra.
Palabras claves: cambios ambientales, detritus, biomasa microbiana, isótopos estables.
INTRODUCTION
Recent studies have demonstrated the importance of detritus in aquatic environments as a source that contributes to metabolic processes of these ecosystems, and the relevance of organic matter dynamics and microbial influence to predict future changes on the energy stability in food chains (Kominoski and Rosemond, 2012).
In aquatic environments, the food chain relationships frequently are maintained by different sources of detritus, due to the large input of allochthonous matter, as also to the decay of aquatic macrophytes (Vannote et al., 1980). Several authors have considered the detritus food chain as the main source maintaining the fish biomass in tropical and subtropical aquatic environments (Araujo-Lima et al., 1986, Benedito-Cecilio et al., 2000, Benedito-Cecilio and Araujo-Lima, 2002, Manetta et al., 2003). In this way, up to 80 % of fish biomass in floodplains is made up by detritivorous species, i.e., dependent on sources of detritus (Agostinho and Zalewski, 1996). However, environmental changes such as modifications in biogeochemical cycles, climate, human activities and changes in the terrestrial ve-getation may affect the amount of detritus and the quality of water, organic matter, and nutrients of these ecosystems, which is reflected in the structure of aquatic webs (Kominoski and Rosemond, 2012).
Floodplains represent areas with high input of organic matter from several origins, and are highly dynamic environments, with well-defined terrestrial, aquatic and transitional environments (Junk et al., 1989). However, Agostinho and Zalewski (1996), Agostinho et al. (2000) and Agostinho et al., (2004) stated that the environmental quality of these areas has experienced a sharp decrease due to deforestation, agriculture, livestock, construction of dams, sand mining and activities of leisure and fishing.
For Agostinho et al., (2000) and Agostinho et al., (2004) a major impact on these ecosystems is the hydrological control by upstream reservoirs, which significantly modifies the flood periods, by changing the intensity, duration and timing of these events. With this, fish assemblages have been negatively affected, mainly the migratory species (Gomes and Agostinho, 1997; Agostinho et al., 2004).
Migratory fish species play a key role in artisanal and recreational fishing. Thus, the lack of flooding periods prevents migratory juveniles to reach marginal lakes to spend the first years of life. This has hindered their reproduction and consequently reduced the fish stock in the region (Gomes and Agostinho, 1997).
The migratory species Prochilodus lineatus (Valenciennes, 1836), popularly known as curimba, in Brazil, is a species of commercial interest and detritivorous with essential role in nutrient cycling, working in the clearance of watercourses subjected to organic pollution (Agostinho, 1985) and in general is a good biondicator by being more sensitive to pollutants (Martinez and Cólus, 2002).
Thus, detritivorous and migratory species are harmed both in relation to detritus sources affected and water resources regulated by reservoirs. Given this, although floodplains have a high biodiversity, these areas are under high environmental disturbances, such as deforestation, and construction of reservoirs. In this case are fundamental studies aiming to examine the microbial community of organic matter, both of allochthonous and autochthonous origin, for understand the importance of these resources for detritivorous fish. This can be assessed, by possible variations in detritus in different environments. In this way, it can be investigated which energy sources (food resources derived from allochthonous and autochthonous matter) are essential to preserve the fish stock, and to analyze possible changes in detritus of the soil and sediment in floodplains.
For these reasons, the focus of this review is an investigative work about the sources of energy for detritivorous fish species in the tropical floodplain.
DETRITUS
Decomposition of Organic Matter
In general, the organic detritus can arise from dead plants or animals with microrganisms action (Mann, 1972). For Swift et al., (1979), it is broadly defined through physical and chemical attributes and interactions with organisms as being any kind of non-living organic matter, including different types of tissues of plants and animals, as well as products secreted and excreted by organisms.
The decomposition of organic matter is based on three stages: leaching, conditioning or catabolism, and fragmentation. Leaching is the process with abiotic removal of organic and inorganic compounds from the material to be decomposed by environmental agents (Davis et al., 2006; Davis and Childers, 2007). In this case, aquatic organisms rapidly incorporate the soluble compounds released as dissolved organic matter, enhancing their utilization by microorganisms (Wetzel, 1995).
The conditioning or catabolism consists in the colonization of the organic matter by fungi and bacteria responsible for the mineralization of the decomposing material (Gonçalves et al., 2006). Thus, this stage prepares the organic matter be consumed by the invertebrate organisms, while the detritus fragmentation enlarges the surface susceptible to the action of microorganisms, resulting in the material degradation (Cunha-Santino and Bianchini Jr, 2006).
In this way, the dissolved organic matter from both autochthonous or allochthonous origin is responsible for microbial production, can be autotrophic, heterotrophic and mixotrophic prokaryotes and eukaryotes, which together make up the microbial food chain (Sherr and Sherr, 2008). Pomeroy (1974) and Azam et al., (1983) have proposed the existence of multiple trophic levels (bacteria, flagellate and ciliate) in the microbial community and outlined important implications for the trophic chain and biogeochemical cycles. This trophic structure is known as microbial loop (Azam et al., 1983), which described the role of bacteria in the absorption and recovery of dissolved organic matter and its transfer to higher trophic levels.
In some studies has been reported that only a small part of the bacterial carbon is passed along the food chain (Cole et al., 2006). It is estimated that the total carbon of prokaryotic cells worldwide corresponds to about 60% carbon estimated for plants (Whitman et al., 1998), evidencing the importance and efficiency of microorganisms in cycling and storage of this element in ecosystems. This efficiency is a result of the balance between bacterial production and respiration that varies widely among aquatic ecosystems. In general, only a very small fraction (< 30%) of the organic matter is converted to microbial biomass (Del Giorgio and Cole, 1998).
Autochthonous and Allochthonous Sources
The energy in food chains of aquatic ecosystems depends on sources of organic matter, both from allochthonous and autochthonous origin, and resources from allochthonous origin are considered essential for aquatic communities, because great part of consumers use the soil organic carbon and submerged letter as a food resources, among them bacteria and protozoa (Berggren et al., 2010), zooplankton (Cole et al., 2011), benthic invertebrates (Solomon et al. 2011) and fish (Weidel et al., 2008; Medeiros and Arthington, 2011; Solomon et al., 2011).
Allochthonous resources can stem from vertical movements, i.e. through falling branches, leaves, flowers, fruit, peels and seeds, as from lateral movements comprising the input of organic matter deposited on the soil into aquatic systems through wind, rainfall, flooding, ground movement and other agents (Lamberti and Gregory, 2007). These sources provide a great part of energy and during dry periods frequently represent the main energy source for aquatic ecosystems (Trevisan and Hepp, 2007).
Studies of the feeding of Characiformes have indicated that these are dependent on food derived from riparian vegetation and changes in the vegetation composition and structure can modify the food availability and hence the feeding habit of the species (Dufech et al., 2003). Other researches, have also pointed out that the allochthonous organic matter especially from the soil taken to the water, is an important food resource for fish (Pusey and Artington 2003; Melo et al., 2004; Weidel et al., 2008; Solomon et al., 2011). This occurs especially, in lotic ecosystems, and has been an important research issue for aquatic ecologists since the beginning of the last century. Only the Journal of the North American Benthological Society encompasses over 158 publications on the subject, since its creation in 1986 until 2010 (Tank et al., 2010).
Autochthonous sources of organic matter include phytoplankton and especially aquatic macrophytes, which are very important in the decomposition stage, when there is transfer of carbon and nutrients to upper trophic levels (Cunha-Santino et al., 2008). Numerous studies have shown the importance of autochthonous sources for aquatic communities (Bunn et al., 2006, Medeiros and Arthington, 2011), highlighting those performed in the Orinoco River that showed that phytoplankton and periphyton were the major sources of carbon for invertebrates and fish (Lewis et al., 2001). Likewise, Araújo-Lima et al. (1986) observed that detritivorous fish of the Amazon River floodplain have used detritus derived from phytoplankton production.
When examining the importance of carbon source, from allochthonous and autochthonous origin, for aquatic organisms, it was verified that in certain situations one becomes more important than the other (Table 1.).
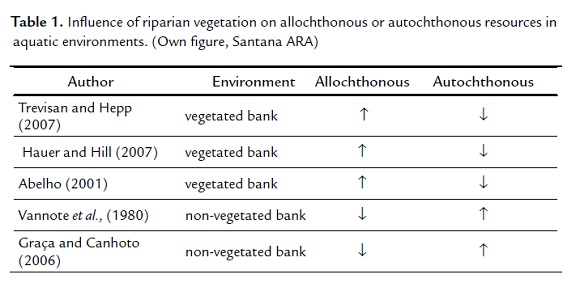
Given this, it is important to take into account physical characteristics of water systems for this analysis. Trevisan and Hepp (2007) mentioned that in rivers with vegetated banks, the allochthonous material is the main energy source for aquatic communities, whereas the autochthonous represents in this case only a small part of energy of rivers with vegetated banks (Abelho, 2001). This is due to the shading of the river by the vegetation that reduces the penetration of solar radiation, reducing the aquatic primary production (Abelho, 2001; Graça and Canhoto, 2006; Hauer and Hill, 2007). However, as the order of the river increases, the input of allochthonous material reduces, prevailing thus the autochthonous primary production (Vannote et al., 1980).
In this way, further studies are required to clarify under what conditions the input of organic matter depends on the allochthonous or autochthonous source, aiming to not only delineate their influence on the carbon cycle of aquatic food chains but also to protect, preserve and implement the biomass of aquatic organisms.
Perspective on the Allochthonous Input in Aquatic Ecosystems
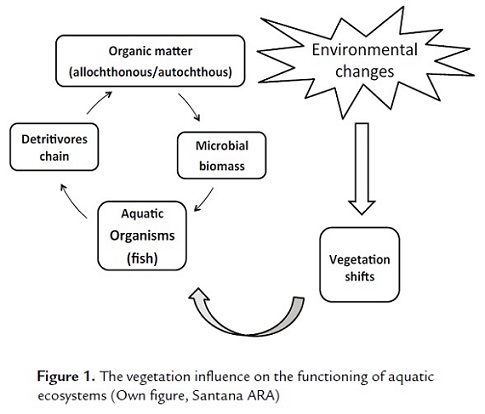
Global environmental changes produce modifications in terrestrial vegetation, which affect diverse aquatic processes because the adjacent vegetation has direct influence on the functioning of these ecosystems (Fig. 1) (Kominoski and Rosemond, 2012).
A basic principle for dynamic changes is the increase in atmospheric CO2 that stimulates the terrestrial primary production and consequently increases the availability of detritus for aquatic ecosystems (Norby et al., 2010). Nevertheless, probably there is also a reduction in the quality of this resource due to the production of secondary compounds (Adams et al., 2003; Adams et al., 2005). These compounds can speed up or slow down the decomposition (Rodrigues, 2006). In this case, the terrestrial primary production can present a greater number of rapid-growth species, predominating vegetation with high photosynthetic efficiency, leading to a significant decrease in the species number, making the ecosystem functionally simple, with few links in trophic chains.
Another effect is due to human activities that change the riparian vegetation or the channel morphology. These activities increase the atmospheric CO2 that affect the aquatic biota owing the high transport of sediment, reduction in allochthonous material, increased temperature and reduced habitat diversity (Poff et al., 2007; Estes et al., 2011).
It has been predicted that food webs of riverine systems will have a low flow rate of permanent organic matter and the detritus quality will be balanced by the carbon lability and amount of nutrients in the ecosystem, environments with high carbon turnover rate will predominate on microbial processes (Kominoski and Rosemond, 2012). Authors still report the importance of future studies to quantify and analyze the basis of the organic matter, and to determine different effects of global changes on allochthonous resources and on microbial processes and adaptations. For this, they predict the importance to associate the community structure with nutrient availability and climatic variations.
In this way, stands out the relevance of flooding areas as ecosystems with significant input of organic matter owing to the inherent feature of seasonal floods. These environments are dynamic and rich in species diversity, and although there are many studies addressing that their preservation also suffer a strong anthropogenic influence, reflected on biotic communities.
Currently there are numerous scientific studies in floodable environments that show the concern on the effect of human activities (Kominoski and Rosemond, 2012). These effects lead to environmental changes that modify the terrestrial vegetation, and promote the increase in atmospheric CO2. This fact influences the dynamics of organic matter and consequently the quantity and quality of allochthonous matter for aquatic communities.
Microbial Biomass of Soil and Sediment
The microbial biomass of soil or sediment is defined as the living fraction of organic matter, excluding plant roots and animal larger than 5 x 10-3μm3, and contributes with 2-5% organic carbon (Jenkinson and Ladd, 1981), besides being an important pool of nutrients for the plants (De-Polli and Guerra, 1996).
In aquatic environments, microorganisms inhabit all surfaces and in low order rivers, the microbial biomass of sediment exceeds the planktonic microbial biomass, but the microbial contribution for the energy flow can change according to seasons (Edwards et al., 1990). For Findlay et al., (2002), in aquatic environments, the accumulation of organic matter in the sediment shelters a high amount of microbial biomass (100-500 mg microbial Cm-2).
The microbial community is known by its ability to decompose organic matter, with a key role in nutrient cycling (Lindemann, 1942; Begon et al., 1996). Meantime, in recent decades it has been assumed that the microbial components play a relevant role in the food webs (Azam et al., 1983, Tranvik, 1989a). Several studies have observed that the biomass and production of microorganisms can be greater than of phytoplankton (Hessen, 1985; Simon et al., 1992; Abreu et al., 1992), and that the carbon flow via microbial trophic interactions can explain the high secondary productivity in ecosystems rich in humic compounds (Edwards, 1987; Tranvik, 1989a).
The participation of microorganisms in the surroundings of aquatic ecosystems in the secondary production can be by up to eight times more than the phytoplankton primary production (Tranvik, 1989b).
The importance of microbiota in aquatic ecosystems is noted, but owing methodological difficulties little is known about microorganisms. However, in recent years ecologists have searched new methods applicable to the microbiota, among them the use of stable isotopes combined with chloroform fumigation-extraction. These tools have been frequently, used in ecology, in the rhizosphere of plants, aiming to estimate and evaluate the microbial biomass (Werth and Kuzyakov, 2009; Werth and Kuzyakov, 2010). Other studies have employed crops and vegetal covers to investigate changes in microbial biomass of soils and to determine how the quality and quantity of vegetation can affect the microorganisms of soil and consequently the carbon turnover (Jin et al., 2010), as well as to examine the impacts caused by preparation systems of soils (Cunha et al., 2012).
QUANTIFICATION OF MICROBIAL BIOMASS IN THE DETRITUS
The use of Stable Isotopes
The conception on the direct contribution of microbial biomass for food chains underwent significant changes over the last 25 years. Nowadays, the question is no longer about the importance of microorganisms in nutrient cycling, but to ask about when, where and why the microbial contribution is so relevant to higher trophic levels (Findlay, 2010).
The first experimental demonstration on the possibility of microorganisms to play a fundamental role in the flow of food webs was developed over 40 years ago by Kaushik and Hynes (1971). The authors observed that the growth of fungi on leaf litter accounted for a significant increase in the quality of this resource. Furthermore, they concluded that the microbial growth in the litter was the main prerequisite for its use by consumers and that the microbial biomass was essentially the only comestible part derived from the leaf litter.
In the same context, Cummins (1974) drew an analogy Peanut butter and crackers when suggested that the microbial biomass is the peanut butter and the substrate leaves are an indigestible cracker. This effective analogy led to a paradigm widely accepted in scientific circles that only microorganisms were available for consumers and the contribution of leaves was insignificant.
Many studies have shown the relationship between the microbial colonization and the nutritional quality of the food (Arsuffi and Suberkropp, 1984; Lawson et al., 1984; Graça et al., 2001). Nevertheless, responses related to the microbial importance vs. the substrate, the leaves in this case, became more complex. Once estimated the microbial biomass, it was possible to determine this relationship (Findlay et al., 1986). For Methvin and Suberkropp (2003), the microbial biomass carbon in the detritus was only a small percentage of the leaf mass. Thus, the absolute mass of the substrate is much greater than the mass of the microbial carbon, in this case, the leaf mass contributed more as food for the consumer. However, the assimilation efficiency of the microbial carbon cold be higher than that obtaining through the plant substrate.
The microbiota play a clear relevant role in the alteration of the nutritional quality of the detritus, generating several ecological effects, besides its contribution to the carbon requirement by consumers. Aquatic hyphomycetes fungi, for example, present different capacities to degrade polymers in leaves (Arsuffi and Suberkropp, 1984). Moreover, they can raise the palatability of submerged plant substrate for detritivorous organisms (Bärlocher, 1992).
This basic observation that consumers of leaf detritus prefer attached microorganisms was confirmed in several studies. Thus, detritus with high microbial biomass can be considered as a resource with high nutritional values not only as storage of microbial carbon (Findlay, 2010).
Studies showed that benthic algae could disproportionately contribute to upper trophic le vels (McCutchan and Lewis, 2002). This study was conducted with stable isotopes, consi dered a fundamental tool to measure the relative contribution of resources, both from allochthonous and autochthonous origin, which maintain the aquatic food chain and could not be evaluated only from the carbon of the organic matter (Carpenter et al., 2005).
Then, a safe way to identify the importance of these items is to follow the matter flow in food chains using stable isotopes (Kling et al., 1992; France and Peters, 1997), which considers that the stable isotope 13C is passed along the food chain, indicating that the consu mer isotopically reflects its diet (DeNiro and Epstein, 1978; Martinelli et al., 1988).
Current isotopic studies have discussed about what carbon source, autochthonous or allochthonous, has contributed more for the feeding of the American detritivorous fish Dorosoma cepedianum (Lesueur, 1818), popularly known as gizzard shad. The authors observed that the autochthonous resources were responsible for most of production of the detritivorous (Babler et al., 2011). Researchers comment that this species uses both food sources, but at different proportions. The study verified that Dorosoma cepedianum uses more than 50% autochthonous resources, and approximately 15% of its production is maintained by allochthonous sources. In other studies, the preferential item of this fish species was detritus with high concentration of carbon and nutrients, and indirectly ingested particles including detritus from terrestrial plants and directly ingested algae and heterotrophic bacteria (Smoot and Findlay, 2010).
On the other hand, Zeug and Winemiller (2008), using 13C isotopic signatures, have shown that allochthonous detritus were responsible for over 80% biomass of Dorosoma cepedianum in main channels of the Brazos River, and both autochthonous and allochthonous resources have similarly contributed in the floodplain system of this area. The authors related this result to the hydrology, because according to Wetzel (1990), the proportion of terrestrial inputs is more intense in floodable areas, while the autochthonous production is higher in rivers.
Given these controversies, the authors suggest that future studies should employ an eco-hydrological approach that quantifies the magnitude of these subsidies, as well as the access routes to aquatic consumers, in different sizes of watersheds and land uses.
The Extraction of Microbial Carbon
Among methods combined with stable isotopes is the chloroform fumigation-extraction, which consists in the quantification of microbial biomass of soils and sediments. Vance et al., (1987) affirmed that it is possible to estimate the carbon contained in the microbial biomass and mentioned that it is proportional to the increase of organic carbon extracted from the soil after chloroform fumigation.
The determination of microbial biomass of soil or sediment is not an estimate of microorganism activity but of the total living microbial biomass of the soil, considered an indicator sensitive to the changes in the organic matter storage (Sparling and Ross, 1993; Matsuoka et al., 2003). This occurs especially in environments under human influence (Insam and Domsch, 1988; Matsuoka et al., 2003).
The evaluation of the microbial biomass of soil is considered an ecological parameter, because it obtains rapid information on the organic changes of soil and allows detecting changes caused by cultivations or deforestations, and measures the soil restoration after removing the surface layer, and evaluated the effects of pollutants (Frighetto, 2000). Moreover, in the floodplain, it is essential to know the origin of carbon sources, because these data can contribute with information about the resources that compose the organic matter and detritus available to food chains. Among the studies on the microbial biomass of soil and sediment, we highlight the use of δ13C, the fact that the bodies reflect, isotopically, its carbon source (DeNiro and Epstein, 1978; Peterson and Fry, 1987). Thus, the composition of the microbiota δ13C indicates the origin of the carbon assimilated by it, revealing the origin of the sources that make up the substrate for microbial production (Peterson and Fry 1987; Pelz et al., 1998; Boschker and Middelburg, 2002), and therefore, the detritus that will be used by detritivorous fish.
The quantification of the percentage of microbial carbon (Cmic) relative to the total organic carbon allows monitoring the disturbances caused by the ecological imbalance, through variations in organic matter. When this ratio increases or decreases rapidly, it is observed instability of the organic matter, may be an increase or a reduction of organic matter of the soil. On the other hand, with a constant ratio, a new balance is reached by this ecological system (Anderson and Domsch, 1989).
Thus the microbial biomass of soil or sediment can be estimated by the concentration of the microbial carbon (Cmic) which is calculated as the difference of the carbon extracted (CF) from the fumigated soil and that of the soil not fumigated (CNF), using the following equation (Vance et al., 1987).
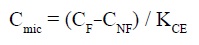
Where for the KCE (extraction coefficient) is used the value of 0.45, according to Wu et al., (1990) and is presented per gram of dry weight.
These tools are widely applied in ecology primarily because the values of microbial carbon and isotopic values of microbial mass enable to identify changes in soils and sediments, long before detecting through physical and chemical alterations (Brookes et al., 1982).
From obtaining the carbon of microbial biomass, it is also possible to determine the isotope value, δ13C, but is necessary to calculate the mass balance according to the equation (Werth and Kuzyakov, 2009)
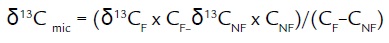
In this way, we can obtain both the value of microbial carbon and, using the stable isotopes of 13C, the isotopic value of microbial biomass, can address the trophic role of microbial carbon which are essential for ecological studies and for monitoring and application of models (Leal and De-Polli, 1999; Turner et al., 2001; Wang et al., 2003; Caraballo et al., 2012).
FINAL CONSIDERATIONS
Considering the above exposed, information about the microbial biomass are fundamental for ecological studies. In recent years, studies at the level of natural microbial communities have revolutionized the microbial ecology and this fact has allowed more accurate identification of organisms that were previously considered as small dots in the microscope.
For many years, ecological studies have assigned to the macrobiota the role of just decomposers of organic matter, but in recent years microorganisms, have been considered, essential for the energy flow in aquatic ecosystems.
Therefore, microorganisms have been the subject of diverse studies. According to the Web of Science® the number of articles in ecology related to microorganisms has increased significantly in the last decades. A research using the combination of the terms microbial biomass and ecology listed 687 articles published between 1990 and 2011.
These data show a significant increase in number of microbial ecological studies. This is especially due to the emergence of new techniques and methods that provided new perspectives in researches on diversity and function of microbial communities in aquatic environments.
Currently, researchers have stated that although challenging, microbial studies are no longer limited by methodological difficulties, and is relevant the inclusion of microorganisms in discussions on ecological subjects. Thus, current and future studies will enable a greater understanding of the role played by microbial processes in aquatic ecosystems.
ACKNOWLEDGEMENTS
The authors are grateful to the Post-Graduate course in Comparative Biology (UEM) and the CAPES for financial support. The Peld (UEM- Nupelia) for bibliographic data as well as to the Institute of Systematic Botany and Ecology, University of Ulm. Germany, for their collaboration.
REFERENCES
Abelho M. From litterfall to breakdown in streams: a review. Sci World J. 2001;1:656-680. Doi:10.1100/tsw.2001.103. [ Links ]
Abreu PC, Biddanda BB, Odebrecht C. Bacterial dynamics of the Patos lagoon estuary, southern of Brazil, relationship with phytoplankton production and suspended material. Estuar Coast Mar Sci.1992;35:621-635. Doi:10.1016/S0272-7714(05)80043-5. [ Links ]
Adams JA, Tuchman NC, Moore PA. Atmospheric CO2 enrichment alters leaf detritus: impacts on foraging decisions of crayfish Orconectes virilis. J N Am Benthol Soc. 2003;22:410-422. [ Links ]
Adams JA, Tuchman NC, Moore PA. Effects of CO2-altered detritus on growth and chemically mediated decisions in crayfish (Procambarus clarkii). J N Am Benthol Soc. 2005;24:330-345. Doi: 10.1899/04-042.1. [ Links ]
Agostinho AA, Zalewski M. A planície alagável do alto rio Paraná: importância e preservação. Maringá, Eduem;1996. 100 p. [ Links ]
Agostinho AA, Thomaz SM, Vera CM, Winemiller KO. Biodiversity in the high Parana river floodplain. In: Gopal B, Junk WJ, Davis JA. Biodiversity in wetlands: assessment, function and conservation. Backhuys Publ, Leiden, Netherlands; 2000. p. 89-118. [ Links ]
Agostinho AA, Gomes LC, Thomaz SM, Hahn NS. The Upper Parana River and its floodplain: main characteristics and perspectives for management and conservation. In: Thomaz SM, Agostinho AA, Hahn NS. The upper Paranáriver and its floodplain: physical aspects, ecology and conservation. Backhuys Publ. Leinden, Netherlands; 2004.p. 381-393. [ Links ]
Agostinho AA. Estrutura da população, idade e crescimento de Rhynelepis abtuse (Osteichthys, Loricariidae) do Rio Paranapanema, Pr. Universidade Federal de São Carlos, São Carlos; 1985. 73 p. [ Links ]
Anderson JPE, Domsch KH. Ratios of microbial biomass carbon to total organic carbon in arable soils. Soil Biol Biochem. 1989;21:471-479. Doi:10.1016/0038-0717(89)90117-X. [ Links ]
Araujo-Lima CARM, Forsberg BR, Victoria R, Martinelli L. Enegy sources for detritivorous fishes in the Amazon. Science. 1986;234:1256-1258. Doi: 10.1126/science.234.4781.1256. [ Links ]
Arsuffi TL, Suberkropp K. Leaf processing capabilities of aquatic hyphomycetes: interspecific differences and influence on shredder feeding preferences, Oikos. 1984;42:144-154. [ Links ]
Azam F, Fenchel T, Field JG, Gray JS, Meyer-Reil LA, Thingstad F. The ecological role of water-column microbes in the sea. Mar Ecol-ProgSer. 1983;10:257-263. Doi:10.3354/meps010257. [ Links ]
Babler AL, Pilati A, Vanni MJ. Terrestrial support of detritivorous fish populations decreases with watershed size. Ecosphere. 2011;2(7):1-23. Doi:10.1890/ES11-00043.1. [ Links ]
Bärlocher F. Research on aquatic hyphomycetes: historical background and overview. In: Bärlocher F. The ecology of aquatic Hyphomycetes. Springer Verlag, Berlin; 1992. p. 1-15. [ Links ]
Begon M, Harper JL, Townsend CR. Ecology. Blackwell Science, Oxford, 1996. [ Links ]
Benedito-Cecilio E, Araújo-Lima CARM. Forsberg BR, Bittencourt MM, Martinelli LA. Carbon sources of Amazonian fisheries. Fisheries Manag Ecol. 2000;7:305-315. [ Links ]
Benedito-Cecilio E, Araújo-Lima CARM. Variation in the carbon isotope composition of Semaprochilodus insignis, a detritivorous fish associated with oligotrophic and eutrophic Amazonian rivers. J Fish Biol. 2002;60:1603-1607. Doi:10.1111/j.1095-8649.2002.tb02453.x. [ Links ]
Berggren M, Strom L, Laudon H, Karlsson J, Jonsson A, Giesler R, et al. Lake secondary production fueled by rapid transfer of low molecular weight organic carbon from terrestrial sources to aquatic consumers. Ecol Lett. 2010;13:870-880. Doi:10.1111/j.1461-0248.2010.01483.x. [ Links ]
Boschker HTS, Middelburg JJ. Stable isotopes and biomarkers in microbial ecology. FEMS Microbiol Ecol. 2002;40:85-95. [ Links ]
Brookes PC, Powlson DS, Jenkinson DS. Measurement of microbial biomass phosphorus in soil. Soil Biol Biochem, Oxford. 1982;14:319-329. Doi:10.1016/0038-0717(82)90001-3. [ Links ]
Bunn SE, BalcombE SR, Davies PM, Fellows CS, Mckenzie-Smith FJ. Aquatic productivity and food webs of desert river ecosystems. In: Kingsford RT. Ecology of Desert Rivers. Cambridge University Press. 2006; p. 76-99. [ Links ]
Caraballo P, Forsberg BR, Leite RG. Papel trófico del microbial loop en un lago de inundación en la Amazonía Central. Acta biol Colomb. 2012;17:103-116. [ Links ]
Carpenter SR, Cole JJ, Pace ML, Van de Bogert MC, Bade DL, Bastviken D, et al. Ecosystem subsidies: Terrestrial support of aquatic food webs from 13C addition to contrasting lakes. Ecology. 2005;86:2737-2750. Doi:10.1890/04-1282. [ Links ]
Cole JJ, Carpenter SR, Kitchell JF, Pace ML, Solomon CT, Weidel B.. Strong evidence for terrestrial support of zooplankton in small lakes based on stable isotopes of carbon, nitrogen and hydrogen. P Natl Acad Sci USA. 2011;108:1975-1980. Doi:10.1073/pnas.1012807108. [ Links ]
Cole JJ, Carpenter SR, Pace ML, Van de Bogert MC, Kitchell JL, Hodgson JR. Differential support of lake food webs by three types of terrestrial organic carbon. Ecol Lett. 2006;9: 558-568. Doi:10.1111/j.1461-0248.2006.00898.x. [ Links ]
Cummins KW. Structure and function of stream ecosystems. Bioscience. 1974;24:631-641. [ Links ]
Cunha EQ, Stone LF, Ferreira EPB, Didonet AD, Moreira JAA. Atributos físicos, químicos e biológicos de solo sob produção orgânica impactados por sistemas de cultivo. Revista Brasileira de Engenharia Agrícola e Ambiental. 2012;16:56-63, Doi: 10.1590/S1415-43662012000100008. [ Links ]
Cunha-Santino MB, Bianchini I. The aerobic and anaerobic decomposition of Typha domingensis Pers. Acta Limnol Bras. 2006;18:321-334. [ Links ]
Cunha-Santino MB, Sciessere L, Bianchini I. As atividades das enzimas na decomposição da matéria orgânica particulada em ambientes aquáticos continentais. Oecol Bras. 2008;12:30-41. [ Links ]
Davis SE, Childers DL. Importance of water source in controlling leaf leaching losses in a dwarf red mangrove (Rhizophora mangle L.) wetland. Estuar Coast Shelf S. 2007;71(1-2):194-201. Doi: 10.1016/j.ecss.2006.07.010. [ Links ]
Davis III Se, Childers DL, Noe GB. The contribution of leaching to the rapid release of nutrients and carbon in the early decay of wetland vegetation. Hydrobiologia. 2006;569:87-97. Doi: 10.1007/s10750-006-0124-1. [ Links ]
Del Giorgio P, Cole JJ. Bacterial growth efficiency in aquatic systems. Annu Rev Ecol Syst. 1998;29:503-541. Doi: 10.1146/annurev.ecolsys.29.1.503 pp.503-541. [ Links ]
De Niro MJ, Epstein S. Carbon isotopic evidence for different feeding partterns in two hyra species occupying the same habitat. Science. 1978;201:916-908. Doi: 10.1126/science.201.4359.906. [ Links ]
De-Polli H, Guerra JGM. Biomassa microbiana: perspectiva para o uso e manejo do solo. In: V.H. Alvarez, L.E.F. Fontes and M.P.F. Fontes. O solo nos grandes domínios morfoclimáticos do Brasil e o desenvolvimento Sustentado. Sociedade Brasileira de Ciência do Solo, Viçosa; 1996. p.551-564. [ Links ]
Dufech APS, Azevedo MA, Fialho CB. Comparative dietary analysis of two populations of Mimagoniates rheocharis (Characidae: Glandulocaudinae) from two streams of Southern Brazil. Neotrop Ichthyol. 2003;1:67-74. Doi: 10.1590/S1679-62252003000100008. [ Links ]
Edwards RT, Meyer JL, Findlay SEG. The relative contributions of benthic and suspended bacteria to system biomass, production, and metabolism in a low gradient blackwater river. J N Am Benthol Soc. 1990;9:216-228. [ Links ]
Edwards RT, Sestonic bacteria as a food source for filtering invertebrates in two southeastern rivers. Limnol Oceanogr. 1987;32:221-234. [ Links ]
Estes JA, Terborgh J, Brashares JS, Power ME, Berger J, Bond WJ, et al. Trophic downgrading of planet Earth. Science. 2011;333:301-306. Doi:10.1126/science.1205106. [ Links ]
Findlay S, Meyer J, Smith PJ. Incorporation of microbial biomass by Peltoperla sp. (Plecoptera) and Tipula sp. (Diptera). J N Am Benthol Soc. 1986;5:306-310. [ Links ]
Findlay EGS, Dye S, Kuehn KA. Microbial growth and nitrogen retention in litter of Phragmites australis compared to Typha angustifolia. Wetlands. 2002;22:616-625. Doi: 10.1672/0277-5212(2002)022[0616:MGANRI]2.0.CO;2. [ Links ]
Findlay S. Stream microbial ecology. J N Am Benthol Soc. 2010;29:170-181. Doi: 10.1899/09-023.1. [ Links ]
France RL, Peters RH. Ecosystem differences in the trophic enrichment of 13C in aquatic food webs. Can J Fishe Aquat Sci. 1997;54:1255-1258. Doi: 10.1139/f97-044. [ Links ]
Frighetto RTS. Análise da biomassa microbiana em carbono: método de fumigação extração. In: Frighetto RTS, Valarini PJ. Indicadores biológicos e bioquímicos da qualidade do solo. Embrapa Meio Ambiente; 2000. p. 157-166. [ Links ]
Gomes LC, Agostinho AA. Influence of the flooding regime on the nutritional state and juvenile recruitment of the curimba, Prochilodus scrofa, Steindacher in upper Paranáriver, Brazil. Fisheries Manag Ecol. 1997;4:263-274. Doi: 10.1046/j.1365-2400.1997.00119.x. [ Links ]
Gonçalves JFJR, França JS, Medeiros AO, Rosa CA, Callisto M. Leaf breakdown in a tropical stream. Int Rev Hydrobiol. 2006;91:164-177. Doi: 10.1002/iroh.200510826. [ Links ]
Graça MAS, Ferreira RCF, Coimbra CN. Litter processing along a stream gradient: the role of invertebrates and decomposers. J N Am Benthol Soc. 200;20:408-420. [ Links ]
Graça MAS, Canhoto C. Leaf litter processing in low order streams. Limnetica. 2006;25:1-10. [ Links ]
Hauer FR, Hill WR. Temperature, light, and oxygen. In: Hauer FR, Lamberti GA. Methods in stream ecology. Second Edition. Academic Press, San Diego; 2007. p. 103-117. [ Links ]
Hessen O. The relation between bacterial carbon and dissolved humic compounds in humic oligotrophic lakes. Microbial Ecol. 1985;31:215-223. Doi: 10.1111/j.1574-6968.1985.tb01152.x. [ Links ]
Insam H, Domsch KH. Relationship between soil organic carbon and microbial biomass on chronosequences of reclamation sites. Microbial Ecol. 1988;15:177-188. Doi: 10.1007/BF02011711. [ Links ]
Jenkinson DS, Ladd JN. Microbial biomass in soil: Measurement and Turnover. In: Paul EA, Ladd JN. New York: Soil Biol Biochem; 1981. p. 415-471. [ Links ]
Jin H, Sun OJ, Liu J. Changes in soil microbial biomass and community structure with addition of contrasting types of plant litter in a semiarid grassland ecosystem. J Plant Ecol. 2010;3:209-217. [ Links ]
Junk WJ, Bayley PB, Sparks RE. The flood pulse concept in river-floodplain systems. In: Doge DP. Proceedings of the International Large River Symposium, Canada: Can Spec Publ Fish Aquat Sci. 1989;106:110-127. [ Links ]
Kaushik NK, Hynes HBN. The fate of dead leaves that fall into streams. Archiv für Hydrobiologie. 197;68:465-515. [ Links ]
Kominoski JS, Rosemond AD. Conservation from the bottom up: forecasting effects of global change on dynamics of organic matter and management needs for river networks. Freshw Sci. 2012;31:51-68. Doi: 10.1899/10-160.1. [ Links ]
Kling WG, Fry B, O'brien WJ. Stable isotopes and planktonic trophic structure in arctic lakes. Ecology. 1992;73:561-566. Doi: 10.2307/1940762. [ Links ]
Lamberti GA, Gregory SV. CPOM transport, retention, and measurement. In: Hauer FR, Lamberti GA. Methods in stream ecology. Second Edition. San Diego: Academic Press; 2007. p. 273-289. [ Links ]
Lawson DL, Klug MJ, Merritt RW. The influence of the physical, chemical, and microbiological characteristics of decomposing leaves on the growth of the detritivore Tipula abdominalis (Diptera: Tipulidae). Can J Zool. 1984; 62:2339-234., Doi: 10.1139/z84-342. [ Links ]
Leal MAA, De-Polli H. Aplicação de modelos ao estudo da matéria orgânica. In: Santos GA, Camargo FAO. Fundamentos da Matéria Orgânica. Porto Alegre: Gênesis; 1999. p. 245-263. [ Links ]
Lewis Jr WM, Hamilton SK, Rodriguez MA, Saunders JF III, Lasi MA. Foodweb analysis of the Orinoco floodplain based on production estimates and stable isotope data. J N Am Benthol Soc. 2001;20:241-254. Doi:10.2307/1468319. [ Links ]
Lindeman RL. The trophic dynamic aspect of ecology. Ecology. 1942;23:399-418. www.jstor.org/stable/1930126. [ Links ]
Manetta GI, Benedito-Cecilio E, Martinelli LA. Carbon sources and trophic position of the main species of fishes of Baía River, Paranáriver floodplain, Brazil. Baz J Biol. 2003; 63:283-290. Doi: 10.1590/S1519-69842003000200013. [ Links ]
Mann KH. Introductory remarks. In detritus and its role in aquatic ecosystems. Proc. IBP-Unesco symposium, Pallanza, Italy. 1972;29:13-16. [ Links ]
Martinelli LA, Matsul E, Forsberg BR, Mozeto AA. Utilização das variações naturais de δ13C no estudo de cadeis alimentares em ambientes aquáticos: princípios e perspectivas. Acta Limnol Bras. 1988;1:859-882. [ Links ]
Martinez CBR, Cólus IMS. Bioindicadores em peixes neotropicais para o monitoramento da poluição aquática na bacia do Rio Tibagi. In: Medri ME, Bianchini E, Shibatta OA, Pimenta JA. A bacia do rio Tibagi. Londrina; 2002. p. 551-557. [ Links ]
Matsuoka M, Mendes IC, Loureiro MF. Biomassa microbiana e atividade enzimática em solos sob vegetação nativa e sistemas agrícolas anuais e perenes na região de Primavera do Leste (MT). Revista Brasileira de Ciência do Solo. 2003;27:425-433. [ Links ]
McCutchan JH, Lewis WM. Relative importance of carbon sources for macroinvertebrates in a Rocky Mountain stream. Limnol Oceanogr. 2002;47:742–752. [ Links ]
Medeiros ESF, Arthington AH. Allochthonous and autochthonous carbon sources for fish in floodplain lagoons of an Australian dryland river. Environ Biol Fish. 2011;90:1-17. Doi: 10.1007/s10641-010-9706-x. [ Links ]
Melo CE, Machado FA, Pinto-Silva V. Feeding habits of fish from a stream in the savanna of Central Brazil, Araguaia basin. Neotrop Ichthyol. 2004;2:37-44. Doi: 10.1590/S1679-62252004000100006. [ Links ]
Methvin BR, Suberkropp K. Annual production of leaf decaying fungi in 2 streams. J N Am Benthol Soc. 2003;22:554-564. [ Links ]
Norby RJ, Warren JM, Iversen CM, Medlyn BE, Mcmurtie RE. CO2 enhancement of forest productivity constrained by limited nitrogen availability. P Natl Acad Sci USA. 2010;107:19368-19373. [ Links ]
Pelz O, Tesar M, Wittich RM, Moore ER, Timmis KN, Abraham WR. Towards elucidation of microbial community metabolic pathways: unraveling the network of carbon sharing in a pollutant-degrading bacterial consortium by immune captureand isotopic ratio mass spectrometry. Appl Environ Microbiol. 1998; 64:4202-4209. [ Links ]
Peterson BJ, Fry B. Stable isotopes in ecosystem studies. Annu Rev Ecol. 1987; 18:293-320. Doi:10.1146/annurev.es.18.110187.001453. [ Links ]
Poff NL, Olden JD, Merritt DM, Pepin DM. Homogenization of regional river dynamics by dams and global biodiversity implications. P Natl Acad Sci USA. 2007;104:5732-5737. [ Links ]
Pomeroy LR. The ocean food web a changing paradigm. Bioscience. 1974;24: 499-504. [ Links ]
Pusey BJ, Arthington AH. Importance of the riparian zone to the conservation and management of freshwater fish: a review. Mar Freshwater Res. 2003;54:1-16. Doi: 10.1071/MF02041. [ Links ]
Rodrigues GG. Decomposição foliar em sistemas ecológicos. In: Mariath JEA, Santos RP. Os avanços da Botânica no início do Século XXI. Sociedade Botânica do Brasil, Porto Alegre: 2006. p. 732-736. [ Links ]
Sherr E, Sherr B. Understanding roles of microbes in marine pelagic food webs: A brief history. In: Kirchman DL. Advances in Microbial Ecology of the Oceans. California: Wiley-Blackwell; 2008. p. 1-24. [ Links ]
Simon M, Cho BC, Azam F. Significance of bacterial biomass in lakes and the ocean: comparison to phytoplankton biomass and biogeochemical implications. Mar Ecol-Prog Ser. 1992;86:103-110. Doi: 10.3354/meps086103. [ Links ]
Smoot JC, Findlay RH. Caloric needs of detritivorous gizzard shad Dorosoma cepedianum are met with sediment bacterial and algal biomass. Aquatic Biol. 2010;8:105-114. Doi: 10.3354/ab00202. [ Links ]
Solomon CT, Carpenter SR, Clayton MK, Cole JJ, Coloso JJ, Pace ML, et al. Terrestrial, benthic and pelagic resource use in lakes: results from a three isotope Bayesian mixing model. Ecology. 2011;92:1115-1125. Doi: 10.1890/10-1185.1. [ Links ]
Sparling GP, Ross DJ. Biochemical methods to estimate soil microbial biomass: Current developments and applications. In: Mulongoy K, Merckx R. Soil organic matter dynamics and sustainability of tropical agriculture. Chichester: John Wiley and Sons; 1993.p. 21-37. [ Links ]
Swift MJ, Heal OW, Anderson JM. Decomposition in Terrestrial Ecosystems. University of California Press, Blackwell Scientific Publications; 1979. 104p. [ Links ]
Tank JL, Rosi-Marshall EJ, Griffiths NA, Entrekin SA, Stephen ML. A review of allochthonous organic matter dynamics and metabolism in streams. J N Am Benthol Soc. 2010; 29:118-146. Doi: 10.1899/08-170.1. [ Links ]
Tranvik LJ. Bacteriplankton in humic lakes: A link between allochtchonous organic matter and pelagic food webs. Lund University; 1989a. 372p. [ Links ]
Tranvik LJ. Bacterioplankton growth, grazing mortality and quantititive relationship to primary production in a humic and a clearwater lake. J Plankton Res. 1989b;11:985-1000. Doi: 10.1093/plankt/11.5.985. [ Links ]
Trevisan A, Hepp LU. Dinâmica de componentes químicos vegetais e fauna associada ao processo de decomposição de espécies arbóreas em um riacho do norte do Rio Grande do Sul, Brasil. Neotrop biol Cons. 2007;2:55-60. [ Links ]
Turner BL, Bristow AW, Haygarth PM. Rapid estimation of microbial biomass in grassland soils by ultra-violet absorbance. Soil Biol Biochem. 2001;33:913-919. Doi: 10.1016/S0038-0717(00)00238-8. [ Links ]
Vance ED, Brookes PC, Jenkinson DS. An extraction method for measuring soil microbial biomass. Soil Biol Biochem. 1987;19:703-707. Doi: 10.1016/0038-0717(87)90052-6. [ Links ]
Vannote RL, Minshall GW, Cummins KW, Sedell JR, Cushing CE. The river continuum concept. Can J Fish Aquat Sci. 1980;37:130-137. [ Links ]
Wang WJ, Dalal RC, Moody PW, Smith CJ. Relationships of soil respiration to microbial biomass, substrate availability and clay content. Soil Biol Biochem. 2003;35:273-284. Doi: 10.1016/S0038-0717(02)00274-2. [ Links ]
Weidel B, Carpenter S, Cole J, Hodgson J, Kitchell J, Pace M, Solomon C. Carbon sources supporting fish growth in a north temperate lake. Aquatic Sci. 2008;70:446-458. [ Links ]
Werth M, Kuzyakov Y. Partitioning of CO2 efflux from soil planted with maize by 13C natural abundance and root exclusion. J Plant Nutr Soil Sci. 2009;172:487-499. Doi: 10.1002/jpln.200700085. [ Links ]
Werth M, Kuzyakov Y. 13C fractionation at the root-microorganisms-soil interface: a review and outlook for partitioning studies. Soil Biol Biochem. 2010;42:1372-1384. Doi: 10.1016/j.soilbio.2010.04.009. [ Links ]
Wetzel RG. Reservoir ecosystems: Conclusions and speculations in Thornton KW, Kimmel BL, Payne FE. Reservoir limnology: Ecological perspectives. John Wiley and Sons. 1990; 227–238. [ Links ]
Wetzel RG. Death, detritus, and energy flow in aquatic ecosystems. Freshw biol. 1995;33:83-89. [ Links ]
Whitman WB, Coleman DC, Wiebe WJ. Prokaryotes: The Unseen Majority. Nat Acad Sci. 1998;95:6578-6583. [ Links ]
Wu J, Joergensen RG, Pommerening B, Chaussod R, Brookes PC. Measurement of soil microbial biomass C by fumigation-extraction: an automated procedure. Soil Biol Biochem. 1990;22:1167-1169. [ Links ]
Zeug SC, Winemiller KO. Evidence supporting the importance of terrestrial carbon in a large-river food web. Ecology. 2008;89:1733-1743. Doi: 10.1890/07-1064.1. [ Links ]
