










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkCES Medicina
versão impressa ISSN 0120-8705
CES Med. vol.28 no.2 Medellín jul./dez. 2014
Artículos de investigación científica o tecnológica
Quantitative EEG analysis disease during resting and memory task in carriers and non-carriers of PS-1 E280A mutation of familial Alzheimer's
Análisis del EEG cuantitativo durante tareas de memoria y reposo en portadores y no portadores de la mutación PS-1 E280A de la enfermedad de Alzheimer
JON EDINSON DUQUE-GRAJALES1, CARLOS TOBÓN2, CLAUDIA P. APONTE-RESTREPO2, JOHN F. OCHOA-GÓMEZ1, CLAUDIA MUÑOZ-ZAPATA2, ALHER M. HERNÁNDEZ VALDIVIESO1, YAKEEL T. QUIROZ-ZAPATA2, FRANCISCO LOPERAN2
1Grupo de investigación en Bioinstrumentación e Ingeniería Clínica (GIBIC), Universidad de Antioquia, Medllín, Colombia
2Grupo de Neurociencias de Antioquia (GNA), Universidad de Antioquia, Medllín, Colombia
ABSTRACT
Background: Alzheimer's disease is the most leading cause of dementia in the world; the mutation PS-1 E280A alters the gene of the Presenilin-1 and causes an early onset familial Alzheimer's disease. This mutation has been found in large kindred of Antioquia, Colombia. The objective of this study was to find differences revealed by electroencephalogram between healthy subjects and asymptomatic carriers that can be used as clinical markers of the disease in this population.Methods: EEG was recorded in 15 asymptomatic E280A carriers and 15 healthy non carriers during resting and a memory task using 64 channels amplifier. Two conditions in the memory task were analyzed: encoding and retrieval, the process of recording and evocating information, respectively. Power spectrum was calculated in delta (0. 5-4. 0 Hz), theta (4. 0-8. 0 Hz), alpha-1 (8. 0-10. 0 Hz), alpha-2 (10. 0-13. 0 Hz), beta (13. 0-25. 0 Hz) and gamma (25. 0-50 Hz) frequency bands for four regions of interest. Changes were evaluated in different conditions by ANOVA analysis..
Results: In resting condition a significant decrease was found in theta (p=0. 0001) and an increase in alpha-2 frequencies (p=0.037) in carriers compare with controls. During encoding of the memory task theta was significantly lower in carriers compared with controls (p=0. 008) and comparing resting versus retrieval process for each group, there was more theta synchronization in carriers.
Conclusions: Early changes in theta frequencies were observed in the EEG recordings, it could be use as clinical markers in this population. Also it seems carriers activate additional cortical regions in order to conserve successful cognitive functions before clinical impairment.
KEY WORDS
Alzheimer's Disease, Quantitative EEG, PS-1 E280A mutation, Theta frequency band, Power spectrum, Memory process.
RESUMEN
Introducción: la enfermedad de Alzheimer es la principal causa de demencia en el mundo; la mutación PS-1 E280A altera el gen presenilin-1 y causa una variante familiar de la enfermedad que se caracteriza por una aparición temprana. La mutación se ha descubierto en un grupo de familias de Antioquia, Colombia. El objetivo de este estudio fue encontrar diferencias, a partir de registros electroencefalográficos de personas portadores de la mutación en una etapa asintomática y sujetos sanos para evaluar si pueden ser utilizadas como un marcador temprano de la enfermedad en la población portadora de la mutación.
Metodología: se realizaron registros EEG en 15 portadores asintomáticos de la mutación E280A y 15 personas sanas no portadoras durante una tarea de memoria y en reposo utilizando un amplificador de 64 canales. En la tarea de memoria se evaluaron dos condiciones: codificación y evocación; el proceso de memorizar y recuperar la información, respectivamente. La potencia espectral fue calculada en las bandas de frecuencia delta (0,5-4,0 Hz), teta (4,0-8,0 Hz), alfa-1 (8,0-10,0 Hz), alfa-2 (10,0-13,0 Hz), beta (13,0-25,0 Hz) y gamma (25,0-50 Hz) para cuatro regiones de interés. Los cambios del espectro fueron evaluados por análisis de varianza ANOVA.
Resultados: bajo la condición de reposo se encontró una disminución importante en la potencia de la banda teta (p=0,0001) y un incremento en la banda alfa-2 (p=0,037) en portadores comparados con controles. Durante la tarea de codificación, los portadores mostraron una disminución significativa en la banda teta (p=0,008). Al comparar reposo contra memoria de evocación se encontró una mayor sincronización en teta en los portadores de la mutación.
Conclusión: se encontraron cambios tempranos de la potencia en la banda teta que pueden ser utilizados como un marcador clínico de la enfermedad en esta población. Una hipótesis adicional basada en los resultados es que los portadores necesitan activar regiones corticales adicionales para conservar las funciones cognitivas antes de empezar un deterioro clínico.
PALABRAS CLAVE
Enfermedad de Alzheimer, EEG cuantitativo, Mutación E280A, Banda de frecuencia teta, Espectro de potencia, Procesamiento de memoria.
INTRODUCTION
Alzheimer`s disease (AD) is the most prevalent cause of dementia, a neurodegenerative con-dition, that generally onset after the age of 65 (1); however there are some mutations that in-duce the onset of the neurocognitive symptoms under this age (2). Until now, three genes have been related to familial Alzheimer's disease: the Presenilin-1 (PS-1) gene, the amyloid precursor protein (APP), and the Presenilin-2 (PS-2) (3,4)
In Colombia, there is a large family group with a mutation in the PS-1 E280A gene, which codifies Presenilin-1, a co-factor of .-secretase involved in the production of b-amyloid (5). This muta-tion has an autosomal dominant inheritance and induces symptoms from the age of 44 years (mild cognitive impairment onset) and dementia at a median age of 46. 8 years (6-9).
Electroencephalography (EEG) might be a powerful and simple tool for the identification of predictive markers of cognitive deterioration, since it can show the rapid and multistage cog-nitive functions that are affected early in neuro-degenerative processes (10).
The main changes in resting EEG recording of sporadic Alzheimer's disease are: slowing of the dominant occipital rhythm (11), early modifica-tions in beta and theta bands and late modifica-tions in alpha and delta bands (12-14). During memory process, one of the earliest and most sensitive EEG changes is the normal synchroni-zation in theta frequencies despite of the increa-se in its relative power (15). EEG records during memory task in sporadic Alzheimer's disease and mild cognitive impairment patients have demonstrated a lack of theta enhancement in the processing of the cognitive stimulus (16-19), demonstrating affectation of the cholinergic system because theta band has been implica-ted in optimal working memory and attentional process (20).
Differences in synchronization of alpha and beta bands also have been reported during memory encoding revealed by synchronization likelihood (a measure similar to coherence but sensible to linear and nonlinear interactions) (21). Syn-chronization likelihood decreased in Alzheimer's disease subjects when they are compared to healthy subjects with subjective memory com-plains and it is related to semantic memory dys-function (22).
EEG studies in carriers of mutations have repor-ted important differences when are compared to non-carriers. EEG recordings in resting of ApoE4 carriers, a genetic risk factor for late-onset spo-radic AD, have shown decrease in alpha and an increase in delta and theta power (23,24). In an Event Related Potential study (ERP) of seman-tic processing in asymptomatic subjects with E280A mutation, lower amplitudes and a diffe-rent topography of N400 potential were obser-ved compared with non-carriers, especially in right inferior-temporal, medial cingulated, left hippocampus and parahippocampus (25).
Another ERP study in the same population showed less positivity in frontal regions and more positivity in occipital regions, during a recognition memory task in asymptomatic ca-rriers compared to controls, which activated frontal areas (26). These differences were more pronounced during the 200-300 msec period.
Finally, a recent quantitative EEG study du-ring resting condition in probable Alzheimer's disease and asymptomatic E280A carriers found alteration in beta frequency and modi-fications in front-temporal regions of the spec-tral parameters before clinical sign of cognitive impairment (3).
In this study we extend the previous research in E280A carriers analyzing the EEG records du-ring resting and a memory task in asymptomatic subjects. The objective was to find differences revealed by EEG between healthy subjects and asymptomatic carriers that can be used as clini-cal markers in this population.
MATERIALS AND METHODS
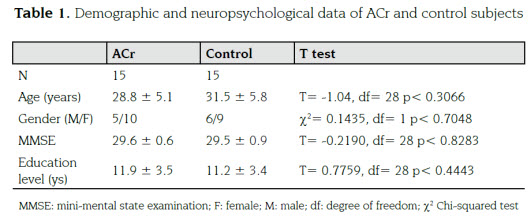
The subjects were members of the E280A mu-tation Colombian kindred. Informed consent for participation was obtained from all subjects ac-cording to a general protocol approved by the Human Subjects Committee of Sede de Investigación Universitaria of University of Antioquia, in Medellin, Colombia. We selected 15 asymptomatic E280A mutation Alzheimer's disease carriers (ACr) and 15 healthy non carriers subjects (control). Table 1 shows the mean values of demographic and clinical characteristic for these groups as well as the results of a T test for each of the variables.
EEG data were recorded with a Neuroscan unit amplifier (Neuroscan Medical System, Neurosoft Inc. Sterling, VA, USA) (0.1 ± 200Hz band pass) from 64 electrodes positioned according to the international 10-10 system with midline refe-rence (subsequently recomputed to common average) at a sample rate of 1000 Hz. A simul-taneous electro-oculogram (0.1 ± 100 Hz band pass) was also recorded. All data were digitized in a continuous recording mode.
Data were registered during an ERP experiment where participants performed a recognition me-mory task using color pictures of concrete and namable objects: 50 new stimuli were presented during the study phase, and 100 stimuli (50% old) were presented during the test phase. Each trial began with a 1,000ms fixation character ("+") prior to the presentation of the stimuli. Study stimuli were then presented for 2,000ms followed by the question, "Do you like this item?" Subjects were then prompted to button press to signify their like/dislike judgment and to remember the items for a subsequent memory test. Test stimuli were presented for 1,500ms, fo-llowed by the question, "Is this item old or new?" Subjects were then prompted to button press to signify their old/new judgment. Subjects were asked to hold their responses until the question appeared immediately after stimuli presentation to minimize response-related ERP artifact (26).
Other recordings were obtained with subjects resting comfortably with their eyes closed du-ring five minutes. Vigilance was continuously monitored in order to avoid drowsiness. For the analysis of the memory EEG data, three condi-tions were selected: encoding, hits and correct rejection. Showing the images for the first time was defined as the encoding condition and it is the process where the participant has to record the new information. Hits were defined as the condition where subjects selected an image as repeated correctly and correct rejection was the condition where subjects selected an image as novel correctly. Both conditions hits and correct rejection are part of retrieval memory process and here the participant has to evocate the lear-ned information. Epochs were selected 200ms before the condition event and 1800ms after. The resting data were segmented in epochs of 2s (2000 samples).
The EEG epochs with ocular, muscular and other types of artifact were removed by a computeri-zed automatic procedure based on linear trend, joint probability and kurtosis approach (27). Then, we used independent component analysis (ICA) approach to identify eye blink, muscle and electrical sources visually detected by an expert (C.T.). These sources were removed of the EEG data. Finally, we repeated the automatic proce-dure in the cleaned EEG ICA-based data to re-move the residual epochs with artifacts.
For quantitative analysis, the fast Fourier trans-form using the Welch approach was computed for 20 artifact-free epochs for each condition (rest, encoding, hits and correct rejection). Re-lative power was calculated in six EEG bands: delta (0. 5-4. 0 Hz), theta (4. 0-8. 0 Hz), alpha-1 (8. 0-10. 0 Hz), alpha-2 (10. 0-13. 0 Hz), beta (13. 0-25. 0 Hz) and gamma (25. 0-50 Hz). The power measures were normalized by dividing the value in one band with the sum of the power in all bands.
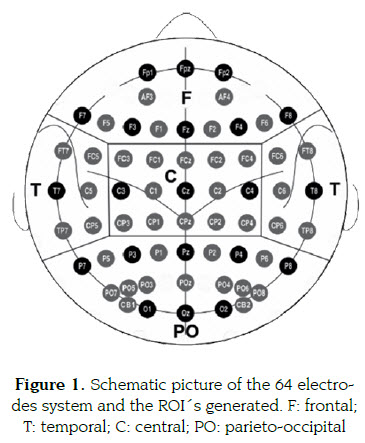
As a result, we had a matrix of 64x6x20 with va-lues of power for each subject, where 64 was the number of channels, 6 the number of bands and 20 the number of epochs selected. Then, four regions of interest (ROI's) were computed: fron-tal -F-: (FP1, FPZ, FP2, AF3, AF4, F7, F5, F3, F1, FZ, F2, F4, F6, F8); temporal -T-: (FT7, FC5, FC6, FT8, T7, C5, C6, T8, TP7, CP5, CP6, TP8); central -C-: (FC3, FC1, FCZ, FC2, FC4, C3, C1, CZ, C2, C4, CP3, CP1, CPZ, CP2, CP4) and parieto-occipital -PO-: (P7, P5, P3, P1, PZ, P2, P4, P6, P8, PO7, PO5, P03, POZ, PO4, PO6, PO8, CB1, O1, OZ, O2, CB2) (figure 1). The original 64x6x20 matrix was reduced to 4x6x20 as the power mean bet-ween the channels of the ROI's for each band. Finally, we averaged the power in the 20 epochs.
The power values were compared by means of ANOVA analysis. The main working hypothesis of the present study was that the power spec-trum in ACr is abnormal compared with control. To test this hypothesis the ANOVA had the fac-tors subjects (ACr and control), ROI's (F, C, T, and PO) and the power spectrum for a band in one condition as independent variable. Also, we compare conditions for each group by means of ANOVA with the factors condition (resting, en-memory task between ACr and control groups as coding, hits and correct rejection), ROI's (F, C, T, and PO) and the power spectrum for a band in one group as independent variable. The statis-tical analysis was carried out in Matlab ®. Sta-tistical significance was defined as the p values lower than 0.05.
RESULTS
Neither demographic information (age, gender and education level) nor neuropsychological examination differences were found (Table 1). No differences were found in the score of the were expected because the asymptomatic con-dition of the population.
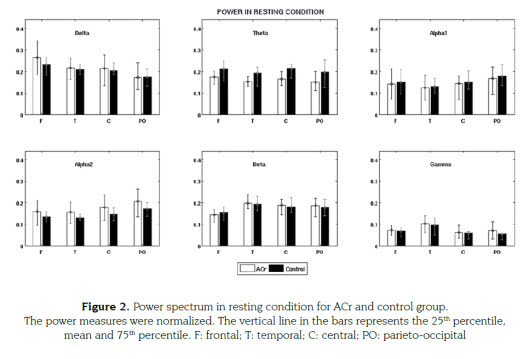
In resting condition the power showed a decrea-se in the theta and alpha-1 bands frequencies for ACr versus control in all ROI's. This de-crease was significant for theta (F=16.52, 1,112 MSe=0.07131; P=0.0001) but not for alpha-1. Also we found a power increase in ACr versus Control for delta and alpha-2 in all ROI's but just the last band was significant (F=4.44, 1,112 MSe=0.0165; P=0.0373) (figure 2). In beta and gamma were not difference found.
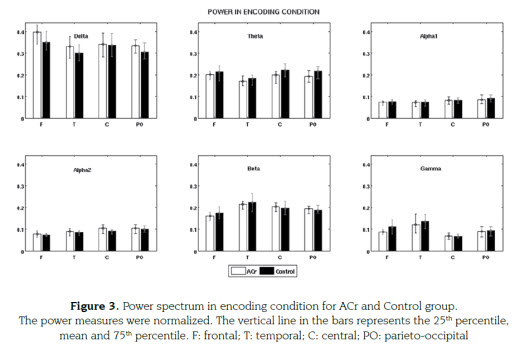
During encoding, theta frequencies were significantly lower in ACr compared with controls (F1,112=7.23, MSe=0,01915; P=0,0083) for all ROIs (figure 3). No differences were found in other bands, neither for hits and correct rejection.
Comparing resting and memory conditions, differences in theta band were found in ACr group for resting versus encoding (F1,112=21.73, MSe=0.0407; P=6.36x10-6), resting versus hits (F1,112=52.13, MSe=0.0654; P=1.98x10-11) and resting versus correct rejection (F1,112=46.38, MSe=0.0589; P=1.86x10-10) for all ROI's. In control group no significant differences were found for resting versus memory. Figure 4 show the magnitude of these powers in both groups.
DISCUSSION
In the present study we analyzed the pattern of the EEG during resting condition and me-mory process revealed by power spectrum. As we expected, comparing the changes of power from resting to encoding there were a decrease of high frequencies (alpha-1, alpha-2, beta and gamma) and an increase in low frequencies (del-ta and theta) for both groups ACr and control. EEG in healthy people during memory process has changes in theta and alpha activity; theta synchronizes with an increased in task demands (theta power increases), whereas alpha desyn-chronizes (alpha power decreases) (28,29).
The activity of theta has been reported in episo-dic long-term and working memory; its oscilla-tions seem to be responsible for the integration of top-down and bottom-up information during the encoding phase, while alpha desynchroni-zation correlates with semantic memory perfor-mance (30,31)
This has been found specially in working me-mory studies, but theta is also associated with attentional processing, spatial navigation and long-term memory processes (18). During me-mory process, we also found a decrease in syn-chronization for alpha frequencies in both, ACr and control, as it has been reported in literature (30,32).
Despite of power in theta frequencies was lower in ACr than control for resting and encoding, if we compare resting versus memory conditions (encoding, hits and correct rejection) for each group, there was more synchronization for all ROI's in ACr. It seems as though ACr subjects create different connectivity pathways in order to get a good cognitive function (compensation hypothesis). This hypothesis has been made through several Alzheimer's disease studies specially in subjects at risk of developing the di-sease because of mutations and this could be observed in early stages of the disease and in preclinical stages (10,26,33,34) like in this study.
There is evidence of neuro-pathologycal changes in carriers of E280A years before onset of symp-toms. Functional imaging during memory task in ACr subjects from 18-26 years old showed grea-ter activation in hippocampal and para-hippo-campal regions and less activation in precuneus and posterior cingulate regions (35). And later at age 28,3 years, there is evidence of accumula-tion of amyloid-beta (Ab) in a florbetapir binding PET study in posterior cingulate, precuneus, pa-rietotemporal, frontal, and basal ganglia regions (36). This early changes could cause a different pattern in the connectivity process and therefo-re in the electrical pattern registered in EEG.
Interestingly, we observed a decreased in theta power frequency during resting condition and memory process at preclinical stages of the di-sease. A similar study in the same population with older asymptomatic carriers (at the age of 39. 9 years) and in probable Alzheimer disease (carriers with mild symptoms) showed a signifi-cant increase of theta during resting condition in probable Alzheimer's disease compared with controls (3). A similar finding had been reported in sporadic Alzheimer's disease (12).
It seems at very early state of the disease (at age of 28. 8 years) there is a decrease in theta band and then it starts to increase at the onset of the disease. It could be explained by the Ab accumulation pattern that begins at age of 28. 3 years. As we said before, there are changes in the activation of brain areas during memory processing in carriers previous to the accumu-lation of Ab and these could be reflected in the decrease of power in theta frequencies revealed by EEG.
As in previous studies in asymptomatic carriers of E280A mutation, we found differences com-pared with controls; there is a decrease in EEG activity in a basal state and affectation of power during memory process mainly in theta frequen-cies. It seems as though carriers need more neural activation in order to have a good neuro-cognitive function as it was revealed by neurop-sychological tests.
CONCLUSION
The EEG recordings revealed early changes mainly in theta frequencies in E280A carriers that can be used as a clinical marker in this po-pulation. Also, it seems these subjects activate additional cortical regions in order to conserve successful cognitive functions before clinical im-pairment; however, more studies are needed in this population in order to clarify the structural and functional difference that these subjects have and that were reveled in the EEG recording but not in the clinical evaluation.
ACKNOWLEDGMENT
Authors thanks to Departamento Administrati-vo de Ciencia, Tecnología e Innovación - Col-ciencias - for the financial support to the Pro-ject "Marcadores neurofisiológicos del inicio de la disfunción cerebral en la etapa pre-clínica de la enfermedad de Alzheimer" identified with the code 1115-519-29028 and to Vicerrectoría de In-vestigación of Universidad de Antioquia (CODI) for financial support to the Project "Identificación de marcadores preclínicos de la mutación E280A de la enfermedad de Alzheimer a partir de medidas de conectividad en EEG" identified with the code PRG14-1-02.
Conflict of interest
There is not conflict of interest in this study.
REFERENCES
1. López OL, Dekosky ST. Clinical symptoms in Alzheimer's disease. Handb Clin Neurol. 2008; 89:207-16. [ Links ]
2. López OL, DeKosky ST. Neuropathology of Alzheimer's disease and mild cognitive im-pairment. Rev Neurol. 16 de julio de 2003; 37 (2):155-63. [ Links ]
3. Rodriguez R, Lopera F, Alvarez A, Fernandez Y, Galan L, Quiroz Y, et al. Spectral Analysis of EEG in Familial Alzheimer's Disease with E280A Presenilin-1 Mutation Gene. Int J Al-zheimers Dis. 2014; 2014:180741. [ Links ]
4. Rogaeva E, Kawarai T, George-Hyslop PS. Genetic complexity of Alzheimer's disease: successes and challenges. J Alzheimers Dis JAD. 2006; 9(3 Suppl):381-7. [ Links ]
5. Van Vickle GD, Esh CL, Kokjohn TA, Patton RL, Kalback WM, Luehrs DC, et al. Preseni-lin-1 280Glu-->Ala mutation alters C-ter-minal APP processing yielding longer abeta peptides: implications for Alzheimer's di-sease. Mol Med Camb Mass. 2008; 14 (3-4):184-94. [ Links ]
6. Duque-Castaño A, Roldán MI, Arango-Viana JC, Arcos-Burgos M, Cubillo H, Lopera F. Neuropathological findings in early-onset Alzheimer's disease (E280a-PS1 mutation). Rev Neurol. 1999; 29 (1):1-6. [ Links ]
7. Lopera F, Ardilla A, Martínez A, Madrigal L, Arango-Viana JC, Lemere CA, et al. Clinical features of early-onset Alzheimer disease in a large kindred with an E280A presenilin-1 mutation. JAMA J Am Med Assoc. 1997; 277 (10):793-9. [ Links ]
8. Acosta-Baena N, Sepulveda-Falla D, Lopera-Gómez CM, Jaramillo-Elorza MC, Moreno S, Aguirre-Acevedo DC, et al. Pre-dementia clinical stages in presenilin 1 E280A familial early-onset Alzheimer's disease: A retros-pective cohort study. Lancet Neurol. 2011; 10 (3):213-20. [ Links ]
9. Ardila A, Lopera F, Rosselli M, Moreno S, Madrigal L, Arango-Lasprilla JC, et al. Neu-ropsychological profile of a large kindred with familial Alzheimer's disease caused by the E280A single presenilin-1 mutation. Arch Clin Neuropsychol Off J Natl Acad Neu-ropsychol. 2000; 15 (6):515-28. [ Links ]
10. Micanovic C, Pal S. The diagnostic utility of EEG in early-onset dementia: a systematic review of the literature with narrative analy-sis. J Neural Transm Vienna Austria 1996. 2014; 121(1):59-69. [ Links ]
11. Kwak YT. Quantitative EEG findings in di-fferent stages of Alzheimer's disease. J Clin Neurophysiol Off Publ Am Electroencepha-logr Soc. 2006; 23 (5):456-61. [ Links ]
12. Baker M, Akrofi K, Schiffer R, Boyle MWO. EEG Patterns in Mild Cognitive Impairment (MCI) Patients. Open Neuroimaging J. 2008; 2:52-5. [ Links ] 13. Jeong J. EEG dynamics in patients with Alzheimer's disease. Clin Neurophysiol. 2004; 115 (7):1490-505. [ Links ]
14. Dauwels J, Vialatte F, Cichocki A. Diagnosis of Alzheimer's disease from EEG signals: where are we standing? Curr Alzheimer Res. 2010; 7(6):487-505. [ Links ]
15. Kim J-S, Lee S-H, Park G, Kim S, Bae S-M, Kim D-W, et al. Clinical implications of quan-titative electroencephalography and current source density in patients with Alzheimer's disease. Brain Topogr. 2012; 25(4):461-74. [ Links ]
16. Basar E, Güntekin B, Tülay E, Yener GG. Evoked and event related coherence of Al-zheimer patients manifest differentiation of sensory-cognitive networks. Brain Res. 2010; 1357:79-90. [ Links ]
17. Güntekin B, Saatçi E, Yener G. Decrease of evoked delta, theta and alpha coherences in Alzheimer patients during a visual oddball paradigm. Brain Res. 2008; 1235:109-16. [ Links ]
18. Karrasch M, Laine M, Rinne JO, Rapinoja P, Sinervä E, Krause CM. Brain oscillatory res-ponses to an auditory-verbal working me-mory task in mild cognitive impairment and Alzheimer's disease. Int J Psychophysiol Off J Int Organ Psychophysiol. 2006; 59 (2):168-78. [ Links ]
19. Yener GG, Güntekin B, Oniz A, Basar E. In-creased frontal phase-locking of event-rela-ted theta oscillations in Alzheimer patients treated with cholinesterase inhibitors. Int J Psychophysiol Off J Int Organ Psychophy-siol. 2007; 64 (1):46-52. [ Links ]
20. Caravaglios G, Castro G, Costanzo E, Di Ma-ria G, Mancuso D, Muscoso EG. . power res-ponses in mild Alzheimer's disease during an auditory oddball paradigm: lack of theta enhancement during stimulus processing. J Neural Transm Vienna Austria 1996. 2010; 117 (10):1195-208. [ Links ]
21. Stam CJ, van Dijk BW. Synchronization like-lihood: an unbiased measure of generali-zed synchronization in multivariate data sets. Phys Nonlinear Phenom. 2002; 163 (3-4):236-51. [ Links ]
22. Pijnenburg YAL, vd Made Y, van Cappellen van Walsum AM, Knol DL, Scheltens P, Stam CJ. EEG synchronization likelihood in mild cognitive impairment and Alzheimer's disea-se during a working memory task. Clin Neuro-physiol. 2004; 115 (6):1332-9. [ Links ]
23. Ponomareva NV, Korovaitseva GI, Rogaev EI. EEG alterations in non-demented indivi-duals related to apolipoprotein E genotype and to risk of Alzheimer disease. Neurobiol Aging. 2008; 29 (6):819-27. [ Links ]
24. De Waal H, Stam CJ, de Haan W, van Straaten ECW, Blankenstein MA, Scheltens P, et al. Alzheimer's disease patients not carrying the apolipoprotein E e4 allele show more severe slowing of oscillatory brain activity. Neurobiol Aging. 2013; 34 (9):2158-63. [ Links ]
25. Bobes MA, García YF, Lopera F, Quiroz YT, Galán L, Vega M, et al. ERP generator ano-malies in presymptomatic carriers of the Alzheimer's disease E280A PS-1 mutation. Hum Brain Mapp. 2010; 31 (2):247-65. [ Links ]
26. Quiroz YT, Ally BA, Celone K, McKeever J, Ruiz-Rizzo AL, Lopera F, et al. Event-related potential markers of brain changes in precli-nical familial Alzheimer disease. Neurology. 2011; 77 (5):469-75. [ Links ]
27. Delorme A, Sejnowski T, Makeig S. Enhan-ced detection of artifacts in EEG data using higher-order statistics and independent component analysis. NeuroImage. 2007; 34 (4):1443-9. [ Links ]
28. Klimesch W, Freunberger R, Sauseng P. Os-cillatory mechanisms of process binding in memory. Neurosci Biobehav Rev. 2010; 34(7):1002-14. [ Links ]
29. Hogan MJ, Swanwick GRJ, Kaiser J, Rowan M, Lawlor B. Memory-related EEG power and coherence reductions in mild Alzheimer's disease. Int J Psychophysiol Off J Int Organ Psychophysiol. 2003; 49 (2):147-63. [ Links ]
30. Klimesch W. EEG alpha and theta oscilla-tions reflect cognitive and memory perfor-mance: a review and analysis. Brain Res Rev. 1999; 29 (2-3):169-95. [ Links ]
31. Sauseng P, Griesmayr B, Freunberger R, Kli-mesch W. Control mechanisms in working memory: a possible function of EEG theta oscillations. Neurosci Biobehav Rev. 2010; 34 (7):1015-22. [ Links ]
32. Basar E, Güntekin B. A short review of alpha activity in cognitive processes and in cogni-tive impairment. Int J Psychophysiol. 2012; 86(1):25-38. [ Links ]
33. Dubovik S, Bouzerda-Wahlen A, Nahum L, Gold G, Schnider A, Guggisberg AG. Adap-tive reorganization of cortical networks in Alzheimer's disease. Clin Neurophysiol Off J Int Fed Clin Neurophysiol. 2013; 12 (1):35-43. [ Links ]
34. Wallace TL, Bertrand D. Importance of the nicotinic acetylcholine receptor system in the prefrontal cortex. Biochem Pharmacol. 2013; 85(12):1713-20. [ Links ]
35. Reiman EM, Quiroz YT, Fleisher AS, Chen K, Velez-Pardo C, Jimenez-Del-Rio M, et al. Brain imaging and fluid biomarker analysis in young adults at genetic risk for autosomal dominant Alzheimer's disease in the prese-nilin 1 E280A kindred: A case-control study. Lancet Neurol. 2012; 11 (12):1048-56. [ Links ]
36. Fleisher AS, Chen K, Quiroz YT, Jakimovich LJ, Gomez MG, Langois CM, et al. Florbe-tapir PET analysis of amyloid-b deposition in the presenilin 1 E280A autosomal domi-nant Alzheimer's disease kindred: a cross-sectional study. Lancet Neurol. 2012; 11 (12):1057-65. [ Links ]
Recibido en: febrero 8 de 2014. Revisado en: mayo 19 de 2014. Aceptado en: mayo 28 de 2014.
