











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introducción
Los sistemas silvopastoriles (ssp) se caracterizan como arreglos agroforestales que incluyen especies arbustivas, arbóreas o maderables, en compañía de un componente herbáceo generalmente asociado a gramíneas (Pezo & Ibrahim, 1998; Sinclair, 1999), que pueden ser desde intensivos (> 10.000 arbustos/ha) sin cobertura arbórea generalizada (Calle et al., 2013), hasta multiestratificados con baja densidad (< 100 árboles/ha) y cobertura arbórea generalizada (Cajas-Giron & Sinclair, 2001).
En el caso de los sistemas multiestratos, las interacciones de facilitación (disminución de la evapotranspiración, fijación de nitrógeno, entre otras) y competencia (agua, luz y nutrientes) entre el componente arbóreo y el herbáceo son elementos importantes en la sostenibilidad y productividad del sistema (Dohn et al., 2013; Guenni, Seiter, & Figueroa, 2008; Martínez, Cajas, León, & Osorio, 2014), especialmente bajo condiciones de pastoreo. Sin embargo, en ocasiones el potencial de productividad herbácea de losssp con cobertura arbórea llega a ser limitado por el efecto de la sombra (Abraham et al., 2014; Paciullo et al., 2017). Usualmente, interceptaciones de radiación incidente por encima del 50 % pueden afectar la producción de biomasa en gramíneas tropicales (Guenni et al., 2008; Paciullo et al., 2017; Villanueva, Ibrahim, & Ríos, 2008), debido a la reducción en la tasa de fotosíntesis, especialmente en especies C4 (Dohn et al., 2013; Santiago-Hernández et al., 2016).
Las interacciones entre la cobertura (dosel) de los árboles y la productividad de las especies herbáceas no son del todo lineales, particularmente en especies de gramíneas que pueden exhibir atributos de plasticidad morfológica y adaptativa (Abraham et al., 2014; Guenni et al., 2008), término que ha sido descrito como la habilidad que poseen algunas plantas para cambiar atributos bioquímicos, morfológicos y fisiológicos, como respuesta ante variaciones ambientales temporales o permanentes (De Kroon, Huber, Stuefer, & Van Groenendael, 2005; Dias- Filho, 2002). Según Guenni et al. (2008), estas adaptaciones en su mayor proporción están asociadas a reducciones en la tasa respiratoria, incremento en la relación parte área/raíz e incremento en el área foliar específica. Otros autores también han citado alongamiento de hojas (Paciullo et al., 2017) y reducción en la tasa de aparición de tallos e incremento en su longitud (Abraham et al., 2014).
En gramíneas (Poaceae) del género Megathyrsus se han reportado evidencias de plasticidad adaptativa bajo condiciones de sombra de árboles, con incremento en atributos morfológicos como altura de la planta, ancho y largo de la hoja, y tasa de aparición de tallos (Medinilla-Salinas et al., 2013; Paciullo et al., 2017; Santiago-Hernández et al., 2016; Silveira, Dos Santos, Rodrigues, Rodrigues, & Alencar, 2017; Viafara, Clavero, & Araujo-Febres, 1997), así como atributos bioquímicos como incrementos en el contenido de proteína cruda (Durr & Rangel, 2003; Paciullo et al., 2017; Santiago-Hernández et al., 2016).
El comportamiento del componente herbáceo, bajo condiciones de cobertura arbórea en un sistema silvopastoril, es parte fundamental de su éxito y adopción. Sin embargo, aún existe poco conocimiento sobre el potencial de las gramíneas en sistemas silvopastoriles con cobertura arbórea y restricción lumínica (Silveira et al., 2017).
El objetivo del presente estudio fue comparar el comportamiento bromatológico y estructural de la gramínea Megathyrsus maximus ( Jacq.) B.K. Simon & S.W.L. Jacobs (Poaceae), bajo cuatro arreglos silvopastoriles de estratos múltiples y una pradera sin árboles.
Materiales y métodos
Área de estudio
La evaluación se llevó a cabo en el centro de investigación Turipaná, vinculado a la Corporación Colombiana de Investigación Agropecuaria (AGROSAVIA), localizado en Cereté, Córdoba, Colombia (08°51' N, 75°49' O, altitud de 18 m s. n. m.), correspondiente a la región Caribe colombiana. La zona presenta dos períodos climáticos definidos: un período de lluvias que se presenta de mayo a noviembre y otro seco entre los meses de diciembre a abril. La precipitación promedio anual es de Colombia (08°51' N, 75°49' O, altitud de 18 m s. n. m.),correspondiente a la región Caribe colombiana.La zona presenta dos períodos climáticos definidos:un período de lluvias que se presenta de mayo anoviembre y otro seco entre los meses de diciembrea abril. La precipitación promedio anual es de1.380 mm, con una temperatura media anual de28 °C. Durante el año, se registra una evapotranspiraciónpotencial de 1.240 mm y una humedadrelativa de 81 % (datos de la estación climatológicaAGROSAVIA-Turipaná). Según la clasificación dezonas de vida de Holdridge (1971), el área se clasificaecológicamente como bosque seco tropical (bs-T).
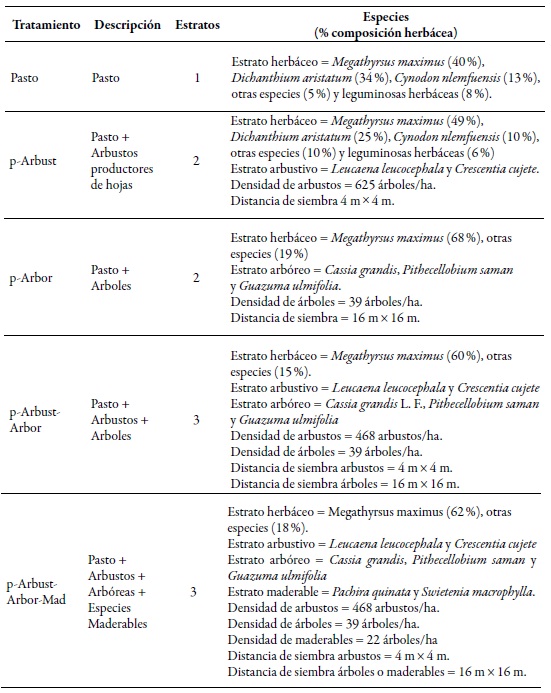
En 1998 se establecieron cuatro tipos de sistemas silvopastoriles de diferente estructura y complejidad, y una pradera sin árboles (control) (Cajas-Giron & Sinclair, 2001). Los arreglos evaluados estuvieron compuestos por tres distintos estratos: pasturas, arbustos y árboles (productores de frutos o maderables). El tratamiento control (Pasto) estuvo conformado solo por pasturas (Cynodon nlemfuensis Vanderyst, Dichanthium aristatum (Poir.) C. E. Hubb. y M. maximus). El primer arreglo silvopastoril presentó solamente arbustos Leucaena leucocephala (Lam.) de Wit. y Crescentia cujete L. (p-Arbust); el segundo incluyó un estrato de árboles Cassia grandis L. F., Pithecellobium saman ( Jacq.) Benth. y Guazuma ulmifolia Lam. (p-Arbor); el tercero (p-Arbust-Arbor), la combinación de pasturas, arbustos y árboles, y el cuarto arreglo (p-ArbustArbor-Mad) incluyó los estratos anteriores más la inclusión de especies maderables Pachira quinata W.S. Alverson y Swietenia macrophylla King (tabla 1).
El experimento se desarrolló en un área de 30 ha, dividida por un efecto diferencial de drenaje en tres sectores caracterizados como bloques (drenaje deficiente, moderado y óptimo). En cada bloque, se estableció una repetición de los tratamientos evaluados en un área de 2 ha (100 m de ancho por 200 m de largo), para un total de tres repeticiones por tratamiento y 15 parcelas (figura 1). Cada tratamiento estuvo dividido en 5 franjas de 4.000 m2, sujetas a un esquema de pastoreo de 2 días de ocupación y 28 días de descanso.
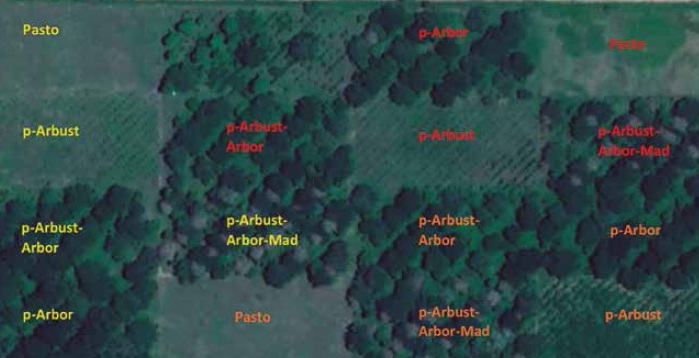
Fuente: Elaboración propia. Imagen satelital de DigitalGlobe® Google 2019. Foto a 1.338 m del suelo.
Figura 1. Imagen satelital de la distribución espacial de los sistemas silvopastoriles en el CI Turipaná (agrosavia), en Cereté (Córdoba, Colombia). Bloque 1: naranja; bloque 2: rojo, y bloque 3: amarillo.
Registro de información
Los cambios bromatológicos y estructurales de la gramínea M. maximus se evaluaron a cuatro edades: 7, 14, 21 y 28 días de rebrote. En cada tratamiento se seleccionaron cuatro puntos centrales para evitar el efecto de borde y recibir las condiciones de sombra propias de cada arreglo silvopastoril. En cada punto (previo corte de uniformización en la gramínea a 25 cm del suelo) se instaló una jaula de exclusión (2 m de largo × 0,5 m ancho; área 1 m2), construida en malla y armadura metálica, para evitar el forrajeo de animales en pastoreo. Cada jaula representó una edad de corte.
En cada una de las parcelas se evaluó la altura de la planta —medida en centímetros, como el promedio de las 5 hojas bandera más altas (Viafara et al., 1997; Wong & Wilson, 1980)—, ancho de la hoja —obtenida en centímetros, de la parte media más ancha de la hoja como el promedio de 5 hojas en la parcela (Guerin, Wen, & Lowe, 2012)—, y relación hoja:tallo (rht) —obtenida a partir de la cosecha total de la parcela y separación manual de hojas y tallos—, para su posterior secado en estufa de ventilación forzada a 60 °C por 48 horas y división del peso seco de las hojas sobre el peso seco de los tallos.
En cada edad, se cosechó una submuestra de forraje para análisis bromatológico. Las muestras de forraje fueron analizadas en el Laboratorio de Nutrición Animal de AGROSAVIA, CI Turipaná, en donde se determinó la proteína cruda (pc), las fibras en detergentes neutro (fdn) y ácido (fda), según el método de la Association of Official Analytical Chemist (AOAC, 2002), y la degradabilidad in situ de la materia seca (dms), según la técnica de la bolsa de nylon descrita por Ørskov, Hovell y Mould (1980). El registro de la información se llevó a cabo durante la época de lluvias (agosto-noviembre del 2010) y seca (enero-abril del 2011), en las cuales se realizaron dos repeticiones cada una.
Análisis estadístico
La información se colectó bajo una estructura de diseño de bloques completos al azar. Cada uno de los registros fue consignado en hojas de cálculo para su revisión y posterior análisis. El análisis de los datos se llevó a cabo considerando un modelo mixto con arreglo de medidas repetidas. Para cada variable, se modelaron los efectos aleatorios mediante un sobre-ajuste de la parte fija del modelo (época, tratamiento y edad). Posterior a la identificación de la mejor estructura de efectos aleatorios (varianza constante del bloque más la varianza aleatoria para cada uno de los niveles del factor edad menos uno), se procedió a la modelación de la parte fija, evaluada desde el modelo que no incluía interacciones (modelo más simple), hasta el modelo que tenía todas las interacciones posibles, a través de estimador de máxima verosimilitud.
Luego de la identificación de la mejor estructura que describía tanto la parte fija como la aleatoria, a través de los criterios de información AIC y BIC (datos no presentados), se procedió al análisis de las variables, con el uso del estimador de máxima verosimilitud restringida (Cayuela, 2018; Di Rienzo, Macchiavelli, & Casanoves, 2009). El análisis de los datos se realizó con la función lme del paquete estadístico nlme (Pinheiro, Bates, DebRoy, & Sarkar, 2018), en el software R-Project (Team R, 2018). En el caso de las variables fda, altura de planta, rht y ancho de la hoja, que no cumplieron los supuestos de normalidad y homogeneidad de varianza, se procedió a la transformación mediante la función logaritmo. Cuando se rechazó la hipótesis nula, se empleó el test de Tukey, para realizar la separación de las medias. En todos los casos se consideró 0,05 como valor crítico.
Resultados y discusión
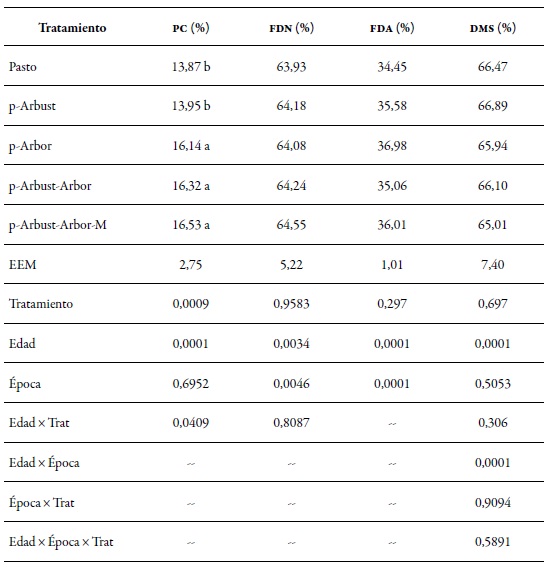
El tratamiento afectó significativamente (p < 0,05) la proporción de pc en la gramínea M. maximus. El promedio de la pc en los tratamientos Pasto y p-Arbust (sin cobertura arbórea) fue 2,42 puntos porcentuales menos que el promedio de los tratamientos p-Arbor, p-Arbust-Arbor y p-ArbustArbor-M (con cobertura arbórea). Para el caso de las fibras, se observó efecto (p < 0,05) de la época sobre las fibras en detergente neutro y ácido. La fdn y la fda para el período seco (65,16 % y 37,30 %, significativamente todas las variables evaluadas respectivamente) fueron mayores a la registrada en (p < 0,05), con aumento para las fibras y disminución el período lluvioso (63,28 % y 37,30 %, respectiva-en la proteína y de la DMS, al pasar de los 7 a los 28 mente). Como era de esperarse, la edad afectó días (tabla 2).
Tabla 2. Composición bromatológica en porcentajes de la gramínea Megathyrsus maximus en cuatro arreglos silvopastoriles y en una pradera sin árboles, en el ci Turipaná, en 2011 pc: proteína cruda; fdn: fibra en detergente neutro; fda: fibra en detergente ácido; dms: digestibilidad de la materia seca;eem: error estándar de la media
Fuente: Elaboración propia
Nota:Letras diferentes entre promedios en la misma columna indican diferencia estadística significativa p < 0,05, según el testde Tukey.
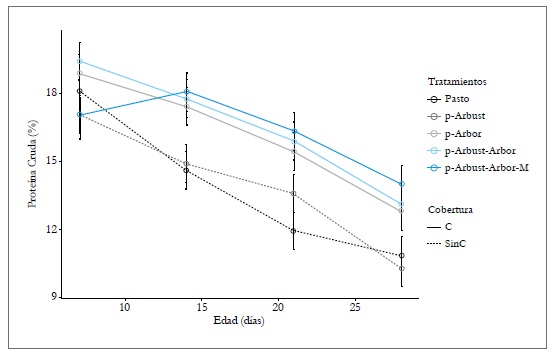
Se observó efecto de la interacción de tratamiento y edad (p < 0,05) para la variable pc. Todos los tratamientos evidencian una tendencia decreciente para el contenido de pc a medida que avanza la edad. Sin embargo, la tendencia es más acentuada en los tratamientos Pasto y p-Arbust (sin cobertura arbórea), comparado con los tratamientos p-Arbor, p-Arbust-Arbor y p-Arbust-Arbor-M (cobertura arbórea). A partir del día 14, los resultados obtenidos para PC difieren significativamente entre el grupo de tratamientos con y sin cobertura arbórea (figura 2).
La tabla 3 ilustra el comportamiento estructural de la gramínea M. maximus, evaluado para los cuatro arreglos silvopastoriles y para la pastura sin árboles. El tratamiento afectó significativamente (p < 0,05) las características altura de la planta, rht y ancho de la hoja. El tratamiento p-Arbor presentó la mayor altura media por planta (80,2 cm), que superó significativamente (p<0,05) la altura media registrada en el tratamiento Pasto (68,8 cm). Este comportamiento fue consistente con la variable ancho de la hoja, en la que se observó que los tratamientos p-Arbor y p-Arbust-Arbor evidenciaron la hoja más ancha (2,68 cm y 2,71 cm, respectivamente) con diferencia significativa del tratamiento Pasto (2,16 cm). Con relación a la variable rht, la tendencia fue contraria a las variables antes descritas. El tratamiento Pasto registró la mejor relación hoja:tallo, con 51,7 g de hojas por cada gramo de tallo. Este registro fue diferente significativamente (p < 0,05) de los registros reportados en los tratamientos con cobertura arbórea p-Arbor (16,6 g), p-Arbust-Arbor (13,6 g) y p-Arbust-Arbor-M (16,5 g).
La época afectó significativamente (p<0,05) las variables altura y ancho de la hoja. La época seca presentó la mayor altura de planta (79,89 cm) y, en promedio, la hoja más ancha (2,61 cm), comparado con el período lluvioso (70,19 cm y 2,41 cm, respectivamente).
Fuente: Elaboración propia
Figura 2. Efecto del arreglo silvopastoril y la edad sobre la proteína cruda en la gramínea Megathyrsus maximus, en CI Turipaná, 2011. C = Cobertura arbórea; SinC = Sin cobertura arbórea. Barras verticales indican el error estándarde la media.
Tabla 3. Características estructurales de la gramínea Megathyrsus maximus en cuatro arreglos silvopastoriles y en una pradera sin árboles, en el ci Turipaná, en 2011 rht: relación hoja: tallo; eem: error estándar de la media.
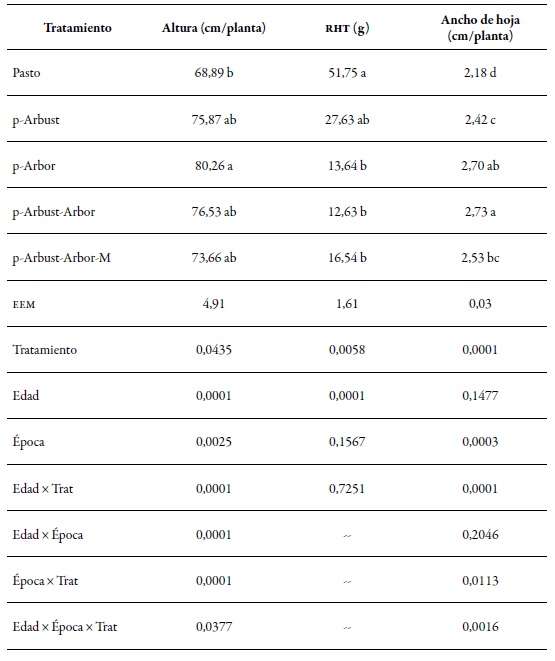
Fuente: Elaboración propia
Nota: Letras diferentes entre promedios en la misma columna indican diferencia estadística significativa p < 0,05 según el test Tukey.
La interacción del arreglo SSP, edad y época presentó efecto significativo (p < 0,05) sobre la altura de la planta y el ancho de la hoja. La altura para el período seco a los 28 días fue mayor en los arreglos SSP p-Arbor (104,77 cm), p-Arbust-Arbor (102,62 cm) y p-Arbust-Arbor-M (111,31 cm), comparado con los tratamientos sin cobertura arbórea (86,39 cm en p-Arbust y 70,97 cm en Pasto). El ancho de la hoja presentó un comportamiento similar a la altura de la planta. Durante el período seco a los 28 días de rebrote, los arreglos ssp con cobertura arbórea registraron los mayores promedios en acho de la hoja (2,80, 2,81 y 2,74 cm para p-ArbustArbor-M, p-Arbust-Arbor y p-Arbor, respectivamente), comparados con el tratamiento Pasto y el ssp p-Arbust, que registraron un ancho medio de la hoja del orden de 2,1 y 2,3 cm, respectivamente (figura 3).
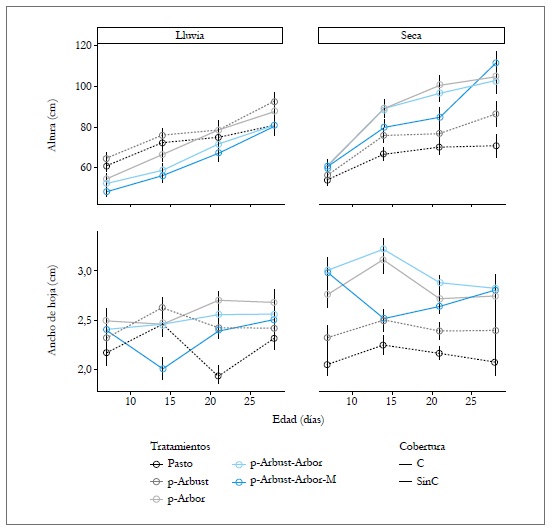
Fuente: Elaboración propia
Figura 3 Efecto del arreglo silvopastoril y de la edad en algunas características estructurales de la gramínea Megathyrsus maximus, en CI Turipaná, 2011. C = Cobertura arbórea, SinC = Sin cobertura arbórea. Barrasverticales indican el error estándar de la media.
Los cambios composicionales y morfológicos observados en M. maximus se ajustan a las descripciones reportadas para especies C4 bajo sombra (Sage & McKown, 2006). Las plantas C4, como la gramínea M. maximus (Giraldo-Cañas, 2010), han evolucionado para adaptarse a ambientes altamente luminosos, calurosos y con baja saturación de CO2, a través de modificaciones bioquímicas y anatómicas que se enfocan en la fijación del carbono a la enzima Rubisco (Edwards & Smith, 2010). Sin embargo, la plasticidad de las C4 a las condiciones de sombra gira en torno a la economía del carbono, en la que la planta incrementa su eficiencia fotosíntetica mediante el aumento del área foliar, la disminución del grosor de las hojas, el cambio la relación parte aérea/raíz y el incremento de la concentración de clorofila, todo ello mediado por la disponibilidad de nitrógeno (Valadares & Niinemets, 2008), que puede ser influenciada por la presencia de árboles leguminosos (Pentón & Blanco, 1997).
En el presente trabajo, los cambios bromatológicos de la gramínea M. maximus bajo condiciones de sombra fueron evidentes, principalmente para la PC, con un diferencial de al menos 2,4 % entre los SSP con cobertura arbórea y los tratamientos p-Arbusto y Pasto. Autores como Paciullo et al. (2017) reportan incrementos del contenido de PC para la gramínea M. maximus del orden de 3,3 % bajo condiciones de sombra artificial (hasta 58 % retención con tela de polipropileno negra); Santiago et al. (2016) registraron incrementos en PC del orden de 2,5 % en hojas y 0,8 % en tallos bajo la cobertura de Melia azedarach L.
Por otra parte, Durr y Rangel (2003) reportaron incrementos 4,7 % en el contenido de PC bajo sombra de árboles de Pithecellobium saman. Contrario a esto, Obispo, Espinoza, Gil, Ovalles y Rodríguez (2008), en la evaluación de niveles de sombra entre 10 % y 30 % generados por P. saman, no reportaron efecto significativo de la sombra en el contenido PC en M. maximus, al igual que Medinilla et al. (2013) bajo cobertura de Gliricidia sepium ( Jacq.) Kunth ex Walp.; estos últimos, probablemente fueron influenciados por la disminución en los contenidos de carbono orgánico en el suelo durante el período experimental.
Con relación al aumento de PC en SSP, Cruz (1997) ha reportado que los incrementos de nitrógeno en gramíneas bajo sombra de árboles están asociados a fenómenos como el reciclaje de nutrientes a través de la caída de hojarascas y a la facilitación de nitrógeno promovida por especies leguminosas. Martínez et al. (2014), evaluando los arreglos SSP objeto del presente estudio, reportaron que, bajo los arreglos con cobertura arbórea (p-Arbor, p-Arbust-Arbor y p-Arbust-Arbor-M), la dinámica de reciclaje de nutrientes mejora los contenidos de P, K, Ca y materia orgánica, comparado con la pradera sin árboles.
Asimismo, autores como Abule, Smit y Snyman (2005), mediante la evaluación de la influencia de los árboles en la composición vegetal y los nutrientes del suelo, han acuñado el término “isla de fertilidad”, para describir las interacciones entre la microfauna del suelo y la acumulación de hojarasca en las mejoras del nitrógeno y la materia orgánica bajo la copa de los árboles. En este sentido, Avendaño-Yanez, López-Ortiz, Perroni y Pérez-Elizalde (2018) determinaron que sistemas naturales de árboles (Vachellia pennatula (Schltdl. & Cham.) Seigler & Ebinger y Lysiloma acapulcense (Kunth) Benth.) y pastos (M. maximus y Hyparrhenia rufa (Nees) Stapf ) incrementan los contenidos de carbono orgánico, materia orgánica, amonio y fósforo total, mediante mecanismos como fijación de nitrógeno, secuestro de nitrógeno y fósforo, por actividad microbiana y a través de la caída y descomposición de la hojarasca.
Estos incrementos de proteína también pueden estar asociados a adaptaciones fisiológicas y bioquímicas evidenciadas en gramíneas bajo sombra. Al respecto, Penton y Blanco (1997) afirman que, bajo condiciones de sombra, se puede presentar una acumulación de nitratos en ausencia de carbohidratos necesarios para su reducción a amoniaco. Esta acumulación de nitratos finalmente conduce al incremento de la participación porcentual de la proteína (N × 6,25) en la materia seca. De igual forma, Dias-Filho (2002) indica que especies de gramíneas C4 son capaces de incrementar significativamente la concentración total de clorofila bajo sombra, comparadas con la misma gramínea a pleno sol. Este incremento de clorofila obedece a la posibilidad de disponer el nitrógeno para la producción de la molécula y de las proteínas ligadas a los pigmentos fotosintéticos (Valadares & Niinemets, 2008), en aras de mejorar la eficiencia en la fotosíntesis.
Los cambios estructurales evidenciados en la altura de la planta, el ancho de la hoja y la rht, influenciados por los arreglos silvopastoriles en la gramínea M. maximus, pueden estar relacionados a la plasticidad morfológica y adaptativa propia de la gramínea, y a los microcambios ambientales (mayor ingreso de luz) que genera el sistema al pasar del período lluvioso al período seco, en el que los árboles (productores de frutos y maderables) experimentan un fenómenos de defoliación total (P. quinata y P. saman) o parcial (C. grandis y G. ulmifolia), como mecanismo de defensa ante la disminución de precipitaciones (Olivero-Lora, 2011; Rusch et al., 2014). El mayor ingreso de luz, producto de la pérdida de hojas de los árboles productores de frutos y maderables en el período seco, pudo estimular la plasticidad morfológica de M. maximus, lo que resultó en mayor incremento de altura media de la planta y ancho de la hoja en los arreglos p-Arbor, p-Arbust-Arbor y p-Arbust-Arbor-M.
La elongación de los tallos en gramíneas bajo condiciones de sombra se considera un mecanismo adaptativo que busca incrementar la capacidad de competencia y mejorar la distribución de la luz a lo largo del dosel (Guenni et al., 2008); de igual forma, el ancho de la hoja refleja una respuesta adaptativa de especies vegetales sometidas a condiciones de estrés (Baruch et al., 2017; Guerin et al., 2012). Paciullo et al. (2017) registraron tasas de elongación de tallo de 0,06 a 0,19 mm/día en M. maximus, al pasar de 0 a 58 % de interceptación de luz. Medinilla et al. (2013) reportaron incrementos de un 12 % de altura en planta para M. maximus, en condiciones de cobertura de G. sepium, comparado con crecimiento a pleno sol. Con relación al ancho de la hoja, Gómez, Navarro y Pérez (2016) indicaron que, bajo cobertura de P. saman, M. maximus registró un incremento significativo de 0,3 cm, comparados con la gramínea a pleno sol.
Conclusiones
Los sistemas silvopastoriles con cobertura arbórea influenciaron una respuesta adaptativa en la gramínea M. maximus, en la proporción de proteína cruda, altura de la planta, ancho de la hoja y relación hoja: tallo. Las respuestas evidenciaron que la gramínea se adaptó a las condiciones de reducción lumínica en los SSP, lo que resulta promisorio en el manejo del pastoreo, con posibles beneficios en el aporte de forraje con mayor calidad proteica a la dieta de los bovinos en pastoreo.
Agradecimientos
Los autores agradecen a AGROSAVIA (anteriormente Corpoica) por el soporte logístico y financiero en el desarrollo de este estudio. También agradecen al Ministerio de Agricultura y Desarrollo Rural, por la financiación del proyecto en el que se desarrolló la presente investigación.
Descargo de responsabilidad
Esta investigación se desarrolló en el marco del proyecto “Valoración Integral de los Beneficios de Sistemas Silvopastoriles de Estratos Múltiples sobre la Sostenibilidad del Suelo y la Productividad Animal en el Sistema Doble Propósito de la Región Caribe de Colombia”, financiado por el Ministerio de Agricultura y Desarrollo Rural en convocatoria 2010, y ejecutado por AGROSAVIA (anteriormente Corpoica).

