










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
versión impresa ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.47 no.2 Santa Marta jul./dic. 2018
https://doi.org/10.25268/bimc.invemar.2018.47.2.748
Research Articles
First record, distribution and ecology of Diplophos proximus (Gonostomatidae, Teleostei) in the Colombian Pacific

1 Investigadora en Ictioplancton y Estudios y Evaluación de Impacto Ambiental. Carrera 47 # 2A - 75 Apto 102 G. Cali, Colombia E-mail: bbeltranleon@yahoo.com.
2 Grupo de Investigación en Biodiversidad Neotropical, Inciva. Avenida Roosevelt # 24 - 80. Cali, Colombia E-mail: raulriosh@gmail.com.
The first findings and distribution of larvae of the species Diplophos proximus, family Gonostomatidae, in the Pacific of Colombia are presented. The analyzes show larvae between Bahía Málaga and Tumaco (limits with Ecuador), with low abundances during the seasons of September 2003 and 2005, July and December 2010, December 2013, February-March 2014, June 2016 and June and September 2017, always collected at stations located more than 113 km away from the coast and around Malpelo island. The larvae are elongated and have noticeable changes during development, initially oval eyes, poorly pigmented body without photophores and finally rounded eyes, dorsal pigments and photophores during transformation. The present work contributes with information about distribution and ecology of larvae of D. proximus becoming the first records of catches for the Colombian Pacific, including the description of the larval stages found.
KEY WORDS: Ichthyoplankton; Gonostomatidae; Deep sea; Colombian Pacific
Se presentan los primeros hallazgos y distribución de larvas de la especie Diplophos proximus, familia Gonostomatidae, en el Pacífico de Colombia. Los análisis muestran larvas entre bahía Málaga y Tumaco (límites con Ecuador), con abundancias bajas durante las épocas de septiembre de 2003 y 2005, julio y diciembre de 2010, diciembre de 2013, febrero-marzo de 2014, junio de 2016 y junio y septiembre de 2017; siempre colectadas en estaciones ubicadas a más de 113 km de distancia de la costa y en los alrededores de la isla de Malpelo. Las larvas son alargadas y tienen notorios cambios durante el desarrollo, inicialmente ojos ovales, cuerpo poco pigmentado y sin fotóforos y finalmente ojos redondeados, pigmentos dorsales y en transformación presentan fotóforos. El presente trabajo contribuye con información de distribución y ecología de larvas de D. proximus, convirtiéndose en los primeros registros para el Pacífico colombiano, incluida la descripción de los estadios larvales encontrados.
PALABRAS CLAVE: Ictioplancton; Larvas; Aguas profundas; Pacífico colombiano
INTRODUCTION
Gonostomatidae is a family of abyssal marine fish commonly known as luminous fish, this family is relatively small and contains only eight genera and thirty- two species. However, the low generic diversity of these fish is compensated by their abundance. Cyclothone, which has twelve species, is considered one of the most abundant vertebrate genera on the planet along with Vinciguerria. These fish are widely distributed in all warm and temperate seas worldwide Atlantic, Indian, and Pacific oceans (Nelson, 2006). Eleven species of gonostomatids are observed in the CalCOFI area (California Cooperative Oceanic Fisheries Investigations), although only four (Cyclothone acclinidens, C. signata, Diplophos taenia and Gonostoma atlanticum) are found with some regularity in the ichthyoplankton samples from the CalCOFI area. Four other species (Cyclothone pseudopallida, Diplophos proximus, Gonostoma ebelingi and G. elongatum) appear mainly offshore south of the CalCOFI stations, and their distributions appear to be largely outside the CalCOFI area (Watson, 1966). Although D. proximus Parr (1931) sometimes appears as synonymous with D. taenia, both species are considered valid according to Ozawa et al. (1990).
Adult gonostomatids are small in size (most are < 7 cm, max 36 cm, usually < 20 cm), have round or ovoid sessile eyes without choroidal tissue, small dentition, and thin branchiostegal rays. The bioluminescence is due to the development of photophores in sizes bigger than 7.5 mm. The fish have intestines of variable length that end at the origin of the anal fin and a generally blackish coloration, although some are silver. These fish are oviparous, and the larvae are elongated and present noticeable changes during development, with initially oval eyes, slightly pigmented bodies without photophores and finally rounded eyes, dorsal pigments and photophores present during transformation. The adults inhabit mainly intermediate depths between 300 and 800 m, and some species rise at night to 50 m from the surface. These fish are found in the epi (0-200 m), meso (200-1000 m) and bathypelagic (> 1000 m) zones. Morphologically, they are very similar to phosichthyids; however, the taxonomic status of these two families has not yet been fully resolved (Watson, 1996).
According to Ozawa et al. (1990), D. proximus presents a large orbital diameter (in proportion to the size of the head, 21-28%) compared to the species D. taenia, D. orientalis and D. australis (15-23%). The diagnostic characteristics are 83-86 total vertebrae (TV), 33-35 abdominal vertebrae (AV) + 49-52 caudal vertebrae (CV), and 92-100 photophores, which is the sum of all ventral rows (IC).
Spawning has not been described for D. proximus, although it has been documented for other gonostomatids, which are oviparous, with eggs and planktonic larvae. The larvae of Diplophos present non-pigmented eyes, elongated yolk sacs and non-functional mouths, and the development is very similar to that of other gonostomatids (Ahlstrom et al., 1984: Ozawa and Oda 1986).
Few larvae and small pelagic juveniles have been found in the collections of CalCOFI from the Gulf of California and near Cabo San Lucas. However, they are much more common in the south, in front to Mexico and Central America. For Colombia, there are no catch records of larvae or adults of this species. Castellanos-Galindo et al. (2006) reported that this species is present in waters adjacent to the Colombian Pacific.
For this Pacific region of Colombia, the following species of gonostomatids can be found: Cyclothone acclinidens, C. microdon, C. signatha, Gonostoma sp. and Yarrella argenteola.Beltrán-León and Ríos (2000) were the first to record the larval stages of C. acclinidens and C. signatha, citing them for subareas III and IV and east of Malpelo Island, with low abundances during the research cruises of April 1993, June-July and December 1995 and November 1998. Therefore, this work contributes distribution information and ecological notes on D. proximus larvae, thus representing the first records of catches for the Colombian Pacific, including descriptions of the larval stages.
STUDY AREA
The Colombian Pacific includes the waters latitudinally between the maritime borders with Costa Rica, Panama and Ecuador and longitudinally from the coastline to the outer limit of the exclusive economic zone created by the island of Malpelo (400 miles from the continental coast). In total, 339500 km2 of water is considered, including estuarine, coastal and oceanic waters, which present depths of up to 4000 m (Beltrán- León and Ríos, 2000).
Located at the western limit of the Eastern Pacific, the Colombian Pacific is characterized by a slight climatic variation that is generated by its location with respect to the sun. Oceanographically, the physical chemistry of the waters depends largely on local phenomena, such as the movement of the Inter Tropical Convergence Zone (ITCZ), continental freshwater drainage, upwelling in Panama, and a complex pattern of surface currents. The main currents that affect the waters of the Colombian Pacific are the cyclonic currents of Panama and Colombia and the equatorial countercurrent.
In general terms, a contrast is observed between oceanic and coastal waters, including estuaries, especially towards the south of Cabo Corrientes. For example, the surface average temperatures vary between 25 and 27°C, and the average salinity is 33.5. Maximum extreme temperatures (up to 31°C) have been detected during El Niño events, while minimums of up to 18°C and salinities close to 35 have been recorded in the first months of the year because of the upwelling of the Gulf of Panama (Prahl et al., 1990). Studies by the Instituto Nacional de Pesca y Acuicultura (INPA) and Zapata et al. (1998) stratified the area into four sub-areas latitudinally from north to south according to the type of coast, width of the platform, contribution of rivers and values of temperature and salinity (Figure 1).
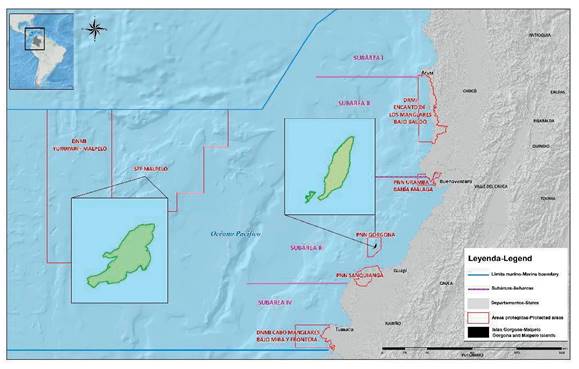
Subarea I: between the border with Panama and Cabo Corrientes (07°13' Lat. N - 05°30' Lat. N). This area is highly influenced by the upwelling of Panama, has high rainfall, little influence of rivers, a narrow platform of approximately 1.8 to 3.7 km and a rocky substrate on the bottom.
Subarea II: between Cabo Corrientes and Málaga Bay (05°30' Lat. N - 04°00' Lat. N). This area has a platform that is wider than the previous and varies between 9 and 19 km, and it receives greater influence from rivers than subarea I, especially due to the large supply of fresh water from the San Juan River, and generally has a muddy sandy substrate on the bottom.
Subarea III: between Málaga Bay and Pasacaballos (04°00' Lat. N - 02°30' Lat. N). This sector has a greater freshwater contribution than the previous two because of the large number of rivers, and its shelf is quite extensive, reaching 56 or 74 km, and it has a sandy mud substrate.
This subarea is characterized as a zone of convergence of coastal flows, whether of oceanic, tidal or fluvial origin. Due to its wide platform and geographical location, water mixing is more evident and longer-lasting than in the other sub-areas of the coast.
In general, the surface arrangement of the oceanographic parameters describes the end-of-year season as seasonal cooling due to the lower surface temperatures (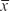 =26°C) (November and December), which is in contrast to the higher temperatures detected at mid-year ( =28°C) (May and July). The surface salinity is lower during the rainy seasons ( =20-29) (May and November), and oxygen is the most homogeneous variable, presenting the highest average value during July (4 ml/l).
=26°C) (November and December), which is in contrast to the higher temperatures detected at mid-year ( =28°C) (May and July). The surface salinity is lower during the rainy seasons ( =20-29) (May and November), and oxygen is the most homogeneous variable, presenting the highest average value during July (4 ml/l).
Subarea IV: located between Pasacaballos and the border with Ecuador (02°30' Lat. N - 01°30' Lat. N). The great contribution of fresh water in this area comes from the Patía and Mira rivers, its platform is very narrow and varies between 9 and 19 km, and it has a sandy muddy substrate.
This area is more influenced by oceanic and equatorial waters (typical of the equatorial countercurrent). For this reason, the oceanographic characterization consists of waters that are colder (27.4-27.6°C) and more hyaline (31-34) than the waters to the north, which are under greater coastal and pluviometric influence. This situation is independent of the annual season, and the intensity of the trade winds allows this countercurrent to reach our coast.
According to Zapata et al. (1998), the importance of the Colombian Pacific is related to its position as a point of confluence of various elements of the underwater fauna of zoogeographic interest that belong to different provinces such as Indo-Pacific, Caribbean, Gulf of California and Peruvian-Chilean, in addition to those of circumtropical distribution. The faunal composition of these waters reflects not only the current dynamics but also the remote geological past, which is still largely unknown.
MATERIALS AND METHODS
The present work was carried out with 24 positive samples for larvae of the species, which were obtained during nine cruises: two ERFEN cruises (“El Niño” Regional Phenomenon Study), which were carried out throughout the Colombian Pacific area in September 2003 and 2005; and seven other cruises performed in a straight line between Buenaventura and the Malpelo Fauna and Flora Sanctuary (including stations around the island) in July and December 2010, December 2013, February-March 2014, June 2016 and June and September 2017. Ichthyoplankton sampling and oceanographic measurements were performed along 10 transects that were perpendicular to the coast, parallel to each other and 54 km apart, with 42 stations for the ERFEN cruises of 2003 and 2005 (Figure 2) and 20 stations for the Buenaventura and the Malpelo Fauna and Flora Sanctuary cruises, with eight in a straight line between Buenaventura and Malpelo and six diurnal and nocturnal stations around the island during 2010 and 2017 (Figure 3).
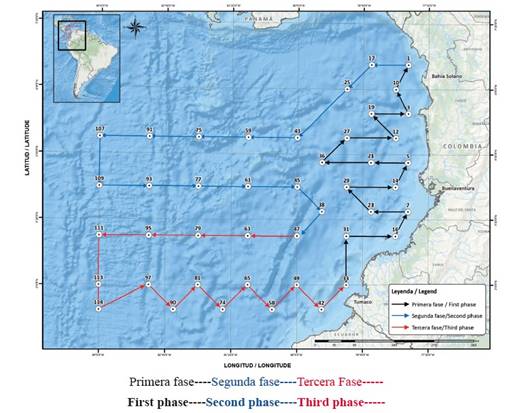
Figure 2 Stations and sampling route of the Cuenca del Pacífico ERFEN cruices 2003 and 2005 (Taken from the CCCP 2004-2007 Cruise Project Execution Plans, ERFEN).
Figure 3 Sampling stations in the Wildlife and Flora Sanctuary Malpelo and the coastal- Malpelo route.
Samples were obtained by a standard bongo net (60 cm diameter), with a mesh of 300 and 500 microns, soft cod-end collectors and Hydrobios flow meters (Figure 4). Oblique trawls were performed according to the methodology established by Smith and Richardson (1979) up to a maximum depth of 200 m. The samples were fixed with formalin neutralized with borax at 10% in seawater. The stations where larvae were captured are shown in Figure 5.

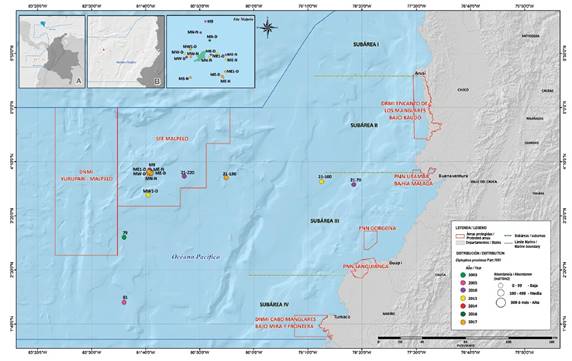
Figure 5 Distribution and abundance of Diplophos proximus larvae collected between 2003 and 2017 in the Colombian Pacific.
The identification of D. proximus larvae was carried out based on the determining meristic, morphometric and pigment characteristics as established in Atlas No. 33 of CalCOFI (Moser, 1996). To determine abundance, the number of larvae and post larvae captured by family, type, genus, and species was considered and the results were standardized to a number of individuals per 10 m2, and for the analysis, the range established by Rueda and Beltrán (1992) was followed: Low: 1 - 99 individuals/10 m2; Medium: 100 - 499 individuals/10 m2; High: > 500 individuals/10 m2. The global analysis of the information allowed drawings to be created with a Wild lucid camera, for each of the stages of the species where the different characteristics associated with a given size are presented.
RESULTS
The collections of D. proximus larvae were carried out during September 2003, September 2005, July and December 2010, December 2013, February-March 2014, June 2016, June and September 2017, in subareas III and IV (Figure 5).
Light fish
Diplophos proximus Parr, 1931 (GONOSTOMATIDAE)
Morphology. PREFLEXION: very elongated body with a preanal length (LpA) 68% of the standard length (LS); head small (<20% LS), eyes rounded and moderately sized (25-33% head length (HL)); mouth large; membrane complete; pectoral fins present but in development. FLEXION: LpA 64% LS; head and eyes small (<25% HL); dorsal, anal, pectoral and caudal fins in development. POSTFLEXION: LpA 62.7% SL; head and eye similar to previous; dentition in both jaws; anal fin developed. TRANSFORMATION: LpA 56-58% LS; head small; only dorsal, anal and caudal fins developed; transverse folds along the large intestine; 36 pre-anal myomeres; 86 total myomeres (Figure 6).
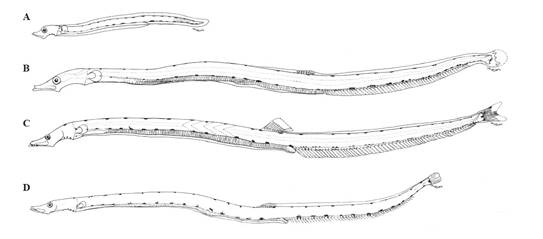
Figure 6 GONOSTOMATIDAE. Diplophos proximus. A. Preflexion 12.7 mm, Station MN-N (1012); B. Flexion 22.5 mm, Station 21-100 (1312); C. Postflexion 26.8 mm, Station 79 (0309); and D. Transformation 45.8 mm, Station MW1-D (1312).
Pigmentation. PREFLEXION: Two or 3 pigments on isthmus in the peduncle of the pectoral fin and over the anus and in rows on the dorsal and ventral edges of the body. FLEXION: In the head, lateral to the brain and in the tip of the jaw. Rows of post-anal pigments in pairs or patches on the ventral edge and base of the rays. Abundant on the caudal peduncle and rays. POSTFLEXION: Similar to the previous, with two over the anus. TRANSFORMATION: Similar to the previous; over the brain and maxilla, large rows of melanophores or patches of dorsal and ventral pigments, with 20 dorsal and 13 ventral (in the tail), (Figure 6, Table 1).
Table 1 Meristics and morphometrics of Diplophos proximus larvae captured in the Colombian Pacific between 2003 and 2017. N: Number of individuals reviewed; V: Number of vertebrae; M: Number of myomeres; D: Dorsal fin; A: Anal fin; Pv: Pelvic fin; Pc: Pectoral fin; C: Caudal fin; Rb: Branchiostegal rays; LT: Total length; LS: Standard length; *F: Fin in formation; Station: 1007 (year, month) ME-N: (Malpelo, East, night) MN-N: (Malpelo, North, night); 21-100: (transect, nautical miles from the coast), ME1-D: (Malpelo, East, 1 mile, day); 79: (ERFEN station); MW-D: (Malpelo, West, day); and MW1-D: (Malpelo, West, 1 mile, day).
| ESTADIO Stage | N | V | M | D | A | Pv | Pc | C | Rb | LT | LS | ESTACIÓN Station |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Preflexión / Preflexion | 3 | 86 | - | - | - | - | - | - | 8.3-12.7 | 8.2-12.6 | ME-N 1007; MN-N 1012 | |
| Flexión / Flexion | 2 | 86 | *F | *F | *F | *F | *F | - | 22.5-28.5 | 22.0-27.7 | 21-100 1213; ME1D 1312 | |
| Postflexión / Postflexion | 2 | 9-10 | 57 | *F | *F | *F | 26.8-28.5 | 26.2-27.2 | 79 0309; ME1-D 1312 | |||
| Transformación / Transformation | 2 | - | 10 | 57-59 | *F | *F | 10+9 | - | 41.3-45.8 | 39.5-44.5 | MW-D 1402; MW1-D 1312 | |
Similar species. The larvae of D. taenia can be distinguished from those of D. proximus by a greater number of anal rays (59-72), myomeres (89-99) and dorsal and ventral pigments.
Habitat. Epi and mesopelagic. Larvae present in subareas III and IV between Malaga Bay and the border with Ecuador between 130 and 463 km from the coast; and to the north, east and west of SFF Malpelo between 0.80 and 1.61 km from the island (Figure 5).
Capture conditions. Surface temperature: 25.3- 28.1°C (X=26.8°C). Surface salinity: 25-34 (X=30.7).
Sampling depth: 1-150 m (X=27.7 m). Distance from the coast: 130-463 km and 0.9 to 1.85 km from Malpelo Island. Time: (03:58 - 20:18), preferably at night and early morning.
Abundance. Low (≤90 ind/m2) in 100% of the stations where it was collected; station 79 (22 ind/10 m2) September 2003; stations 81 (6 ind/10 m2) and M9 (12 ind/10 m2)) in September 2005; stations ME-D (2 ind/10 m2) and ME-N (1 ind/10 m2) in July 2010; stations 21-70 (2 ind/10 m2), 21-220 (1 ind/10 m2) and MN-N (1 ind/10 m2) in December 2010; stations 21-100 (2 ind/10 m2), ME-N (3 ind/10 m2), MN-N (4 ind/10 m2)), ME1-D (2 ind/10 m2), MW-D (4 ind/10 m2) and MW1-D (2 ind/10 m2) December 2013, MW-D station (2 ind/10 m2) from February to March 2014; station MN-D (4 ind/10 m2) in June 2016; stations 21-190 (1 ind/10 m2), MS-N (2 ind/10 m2), ME-D (2 ind/10 m2), Nevera D (2 ind/10 m2) in June 2017; and ME-N stations (3 ind/10 m2), MW-N (4 ind/10 m2), ME1-D (2 ind/10 m2) and MW1-D (4 ind/10 m2) in September 2017. Larvae of this species were not caught on other cruises occurring on dates other than in June, July, September, December and February-March or on cruises made less than 130 km from the coast. The highest frequency and abundance were found during the months of September and December (Figure 5).
Range. The range is circumglobal in tropical and temperate waters, and it also ranges to the tropical eastern Pacific in southern California. This record is the first for the Colombian Pacific. The distribution found for the larvae shows a latitudinal range between Málaga Bay (04° 00' Lat. N) and the border with Ecuador (01°30' Lat. N) in subareas III and IV and a longitudinal range from 130 km from the coast (78º27'26.2 W) to the waters surrounding the SFF Malpelo (Figure 5).
DISCUSSION
The collections of D. proximus larvae were carried out in September 2003, September 2005, July and December 2010, December 2013, February-March 2014, June 2016, June and September 2017, years coinciding with periods after “El Niño” events (i.e., June 2002-March 2003, August 2004-February 2005, August 2009-May 2010, February 2015-June 2016, December 2016-March 2017) and during “La Niña” events (i.e., August 2010-May 2011, July 2016-November 2016, October 2017-February 2018) (Golden Gate Weather Services, 2015). For Colombia, the species was found in post-El Niño periods, normal periods (Dec/13 and Feb-Mar/14) and Niña and post- Niña periods. This pattern is different from that recorded by Funes-Rodríguez et al. (2006) for Baja California Sur, who claim that D. proximus together with Diaphus pacificus and Benthosema panamense were part of a group of tropical species useful to distinguish El Niño event of 1982-84. Similarly, Avendaño-Ibarra et al. (2003), recorded other important tropical demersal and mesopelagic taxa, such as Benthosema panamense, Symphurus williamsi, D. proximus and Cubiceps pauciradiatus, which contributed 12.2% of the total abundance during the fall of 2003 for Baja California Sur and is consistent with previous reports during warm periods with positive sea surface temperature (SST) anomalies (Funes-Rodríguez et al., 1995, 2006).
Ozawa et. al. (1990) reported that D. proximus is valid as endemic in the tropical eastern Pacific, and Fourriére et al. (2016) reported that it occurs in the reefs of the Revillagigedo archipelago and areas nearby Mexico. However, D. proximus is often considered synonymous with D. taenia Günther despite being a different species, and it has thus been described in its larval stage in Atlas No. 33 of CalCOFI (Moser, 1996), this information has not been included in the FishBase database.
CONCLUSIONS
In Colombia, the species D. proximus has been found during normal climatic periods and “El Niño” and “La Niña” periods, whereas in Baja California Sur, the species has been recorded during warm periods and periods with positive temperature anomalies; therefore, the species is likely part of a group of tropical species that can be used to distinguish the “El Niño” event in other latitudes.
Larval stages of D. proximus have been found from 2003 to the present with a relatively wide distribution in the south-central Colombian Pacific; thus, a population of potentially reproductive adults of the species must inhabit this zone.
Because of the lack of captures or catch records of adults of the species of the family Gonostomatidae as well as many other species of other families of meso and bathypelagic habitats, more complete and detailed studies in colombian deep waters should be conducted.
ACKNOWLEDGEMENTS
The authors are grateful to the crews of the M/N “María Patricia” on the cruises to the SFF Malpelo and the B/O ARC Providence of the National Navy on the ERFEN Cruises for their support during the planning and execution of the sampling expeditions; the biologist Yaneth Morales, who participated in the sampling and the separation phase in the laboratory by the ERFEN cruise (September 2005) and SFF Malpelo (July and December 2010); the biologist Maicol Ramírez (of AG Consultores Ambientales S.A.S. (AG Environmental Consultants) for his collaboration in sampling during December 2013; the ecologist Rodrigo Baos (WWF-Colombia) for his collaboration in the sampling during February 2014; the biologists Pablo Jiménez and Graciela Serna (WWF-Colombia) for their collaboration in the sampling during June and September of 2017; WWF-Colombia, who provided financial support (TP 09 Agreement) for the sampling and/or separation phase in the laboratory of the ERFEN cruise of September of 2005; the Malpelo Foundation for financing of the cruises to the SFF Malpelo; the National Parks for providing financial support for the participation of Y. Morales in the cruise to SFF Malpelo in July and December 2010; Gustavo Gutiérrez (AG Consultores Ambientales SAS) for providing financing in the sampling and/or separation phase in the laboratory of the cruise of December 2013; the entities participating in the cruise ships and their respective researchers for providing support; José Tavera (Universidad del Valle) for sending literature; Luis Zapata (WWF-Colombia) for reviewing the manuscript; Arturo Acero (Universidad Nacional de Colombia sede Caribe) for providing support; and Leydi Cuadros (WWF-Colombia) for preparing the maps.
REFERENCES
Ahlstrom, E. H., W. Richards and S. H. Weitzman. 1984. Families Gonostomatidae, Sternoptychidae, and associated stomiiform groups: development and relationships. 184-198. In: Moser, H.G., W. J. Richards, D.M. Cohen, M.P. Fahay, AW. Kendall Jr. and S. L. Richardson (Eds.). Ontogeny and systematics of fishes. Am. Soc. Ichthyol. Herpetol. Spec. Publ., 1. [ Links ]
Avendaño-Ibarra, R., R. De Silva-Dávila, F. Ordóñez-Guillén y G. Vázquez-López. 2003. Composición estacional de larvas de peces frente a Baja California Sur (primavera y otoño de 2003). 413 - 431. En: Gaxiola-Castro G. y R. Durazo (Eds.). Dinámica del ecosistema pelágico frente a Baja California 1997-2007. Diez años de investigaciones mexicanas de la corriente de California. SEMARNAT-INE-CICESE-UABC, México D.F. 501 p. [ Links ]
Beltrán-León, B. S. y R. Ríos. 2000. Estadios tempranos de peces del Pacífico colombiano. Instituto Nacional de Pesca y Acuicultura -INPA- ISBN: 958-9356-03-6. Buenaventura. 727 p. [ Links ]
Castellanos-Galindo, G.A., E.A. Rubio, B.S. Beltrán-León and C. Baldwin. 2006. Checklist of stomiiform, aulopiform and myctophiform fishes of the Tropical Eastern Pacific. Biota Col., 7(2): 245-262. [ Links ]
Centro Control Contaminación del Pacífico - CCCP. 2004-2007. Planes de ejecución cruceros regionales de investigación oceanográfica (ERFEN) en el Pacífico Sudeste 2004 - 2007. Tumaco. 15 p. [ Links ]
Fourriére, M., H. Reyes-Bonilla, A. Ayala-Bocos, J.A. Ketchum and J.C. Chávez-Comparan. 2016. Checklist and analysis of completeness of the reef fish fauna of the Revillagigedo Archipelago, Mexico. Zootaxa, 4150 (4): 436-466. [ Links ]
Fricke, R. W.N. Eschmeyer and R. van der Laan (Eds.). 2018. Catalog of fishes: Genera, species, references. http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp 18/04/2018. [ Links ]
Funes-Rodríguez R, R. González-Armas y A. Esquivel-Herrera. 1995. Distribución y composición específica de larvas de peces durante y después de El Niño, en la costa del Pacífico de Baja California Sur (1983-1985). Hidrobiología, 5: 113-125. [ Links ]
Funes-Rodríguez, R., A. Hinojosa-Medina, G. Aceves-Medina, S.P. Jiménez-Rosenberg and A.J. Bautista-Romero. 2006. Influences of El Niño on assemblages of mesopelagic fish larvae along the Pacific coast of Baja California Sur. Fish. Oceanogr., 15 (3): 244-255. [ Links ]
Golden Gate Weather Services, 2015. Eventos de El Niño y La Niña en el siglo XXI. 23/09/2015. [ Links ]
Moser, G. 1996. The early stages of fishes in the California Current Region. Atlas No. 33. CalCOFI. 1505 p. [ Links ]
Nelson, J. S. 2006. Fishes of the world. Third Edit. Fourth Edition. Wiley, New York. 601 p. [ Links ]
Ozawa, T. and K. Oda. 1986. Early ontogeny and distribution of three species of the gonostomatid genus Diplophos in the western North Pacific. 74-84. In: Ozawa, T . (Ed.). Studies on the oceanic ichthyoplankton in the western North Pacific. Kyushu Univ. Press, Fukuoka. [ Links ]
Ozawa, T ., K. Oda and T. Ida. 1990. Systematics and distribution of the Diplophos taenia species complex (Gonostomatidae), with a description of a new species. Jap. J. Ichthyol., 37(2): 98-115. [ Links ]
Prahl, H. v., J.R. Cantera y R. Contreras. 1990. Manglares y hombres del Pacífico colombiano. Fondo FEN, Bogotá. 193 p. [ Links ]
Rueda-Montenegro, C. y B. S. Beltrán-León. 1992. Ictioplancton de las familias Clupeidae y Engraulidae en el litoral Pacífico colombiano. 741-752. En: Mem. VIII Sem. Nal Cienc. Tecnol. Mar Congr. Centroam. Car. Cienc. Mar, Santa Marta. [ Links ]
Smith, L. y S. Richardson. 1979. Técnicas standard para prospecciones de huevos y larvas de peces pelágicos. FAO. Doc. Tec. Pesca, 175: 107. [ Links ]
Watson, W. 1996. Gonostomatidae: Bristlemouths. 247-267. In: Moser, H.G . (Ed.). The early stages of fishes in the California Current Region. California Cooperative Oceanic Fisheries Investigations (CalCOFI) Atlas No. 33. Allen Press, Inc., Lawrence, Kansas. 1505 p. [ Links ]
Zapata, L. A., G. Rodríguez, B. Beltrán, G. Gómez, W. Angulo, A. Gómez e Y. Morales 1999. Prospección de los principales bancos de pesca en el Pacífico colombiano, durante noviembre de 1998. Bol. Cient. INPA, (6): 111-175 [ Links ]
Received: May 04, 2018; Accepted: August 23, 2018
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
