











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
Lo que se conoce sobre la biología de los peces marinos no puede considerarse suficiente debido al escaso conocimiento sobre la taxonomía y ecología de sus larvas. Ecológicamente, las larvas y los adultos de los peces son diferentes en cuanto a sus requerimientos de hábitat, alimento e incluso varían en cuanto a su comportamiento (Sameoto, 1984, Allen, Pondella & Horn, 2006). En este sentido, ha sido reiteradamente reconocido que la escasa información disponible sobre la taxonomía de las larvas de peces no solo es un obstáculo en el avance teórico, sino que también es una problemática recurrente en los esfuerzos realizados para lograr un adecuado manejo de este importante recurso alimentario (Sale, 1980; Govoni, 2005; Fuiman & Werner, 2009; Brochier et al., 2013).
En términos generales, la investigación en ictioplancton requiere de una mayor dedicación y esfuerzo debido a la problemática que representa su identificación taxonómica (Richards, 1985; Leis, 1991; Hernández et al., 2013; Leis, 2015), al punto que la literatura al respecto es escasa, muchas de las descripciones tienen poca calidad, han sido descritos estadios larvales de pocas especies de peces, se desconocen los estadios de desarrollo larval de una gran cantidad de familias y los esfuerzos científicos en esta temática generalmente han estado direccionados a pocas regiones geográficas o hacia especies de interés comercial (Olney, 1983; Sanvicente-Añorve, Flores-Coto & Chiapas-Carrara, 2000; Duffy‐Anderson et al., 2006; Boeing & Duffy-Anderson, 2008; Auth & Brodeur, 2013). Estas limitaciones son aún más acentuadas en áreas tropicales, en donde la diversidad de peces es más elevada y la abundancia larval, para gran parte de las taxa recolectadas, es baja (Leis, 1993, Franco-Gordo et al., 2003; Hernández et al., 2013).
Para el Pacífico colombiano, los escasos esfuerzos de investigación en ictioplancton han estado direccionados principalmente el análisis de aspectos ecológicos relacionados con especies de interés comercial o la descripción de los ensamblajes de larvas de peces en sitios específicos de los ambientes costeros (Beltrán, Ramos, Escobar & Ríos, 1994; Escarria, Beltrán-León, Giraldo & Zapata, 2006; Escarria, Beltrán-León, Giraldo & Zapata, 2007; Martínez-Aguilar, Giraldo & Rodríguez-Rubio, 2010; Medina-Contreras et al., 2014; Calle-Bonilla, Giraldo & Cuéllar-Chacón, 2017). Sin embargo, se destacan los trabajos sobre la taxonomía de larvas de peces realizado por Ahlsthrom (1971), quien identificó la presencia de larvas de más de 50 familias de peces y el trabajo realizado por Beltrán-León & Ríos (2000), quienes describen las características taxonómicas de las larvas de 236 especies de peces del Pacífico colombiano.
Ha sido ampliamente demostrado que la estructura de los ensamblajes de larvas de peces es altamente variable en cuanto a su composición, respondiendo de manera específica a las variaciones ambientales propias de cada localidad (Whitfield & Pattrick, 2015; Houde, 2008; León-Chavez et al., 2010; De Macedo-Soares et al., 2014; León-Chávez et al., 2015; Keskin & Pauly, 2019). En este sentido, el objetivo de la presente investigación es consolidar la identificación taxonómica de las larvas de peces presentes en Cabo Manglares - DNMI Cabo Manglares, Bajo Mira y Frontera, región sur del Pacifico colombiano, considerando la variación estacional de las condiciones de temperatura y salinidad que han sido descritas para el Pacífico colombiano (Rodriguéz-Rudio & Scheneider, 2003; Devis-Morales, Schneider, Montoya-Sánchez & Rodríguez-Rubio, 2008; Giraldo, Rodríguez-Rubio & Zapata, 2008; Rodríguez-Rubio & Giraldo, 2011; Valencia, Lavaniegas, Giraldo & Rodríguez, 2013; Villegas, Málikov & Díaz, 2016; Jerez-Guerrero, Críales-Hernández & Giraldo, 2017).
MATERIALES Y METODOS
Cabo Manglares se localiza en el sector sur del litoral Pacífico Colombiano (1° 38’ N - 78° 58’ W), en el departamento de Nariño. En 2017, se alinderó y declaró en esta región el Distrito Nacional de Manejo Integrado Cabo Manglares, Bajo Mira y Frontera, como área marino-costera protegida por el Ministerio de Ambiente y Desarrollo Sostenible, abarcando 190.282 Ha de paisaje marino-costero correspondiente al delta del río Mira e incluyendo unidades ecosistémicas de fondos marinos, playas, esteros, manglares y natales (MADS, 2017).
Para obtener las larvas de peces se realizaron pescas de zooplancton en 32 estaciones de muestreo asociadas a la zona costera de Cabo Manglares (figura 1). Con el propósito de incorporar la variación estacional de las condiciones oceanográficas que ha sido descrita para el Pacifico colombiano, se realizaron dos campañas de muestreo, la primera en diciembre de 2016 y la segunda en marzo de 2017. En cada estación, se utilizó una red Bongo de 33 cm de boca, con malla de 300 mm y 500 mm para capturar el zooplancton, mediante arrastres oblicuos de 10 minutos de duración, desde 20 m de profundidad hasta la superficie. Se adosó un flujómetro Hydrobios a cada boca de la red con el propósito de cuantificar el volumen de agua filtrado. Todos los muestreos fueron realizados entre las 08:00 y las 16:00 horas. Las muestras recolectadas fueron preservadas en formalina tamponada en agua de mar al 4 % para su transporte al laboratorio.
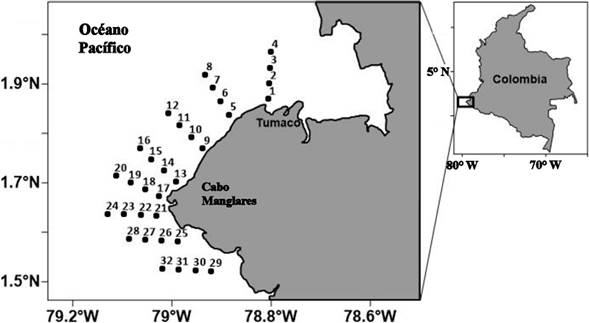
Figura 1 Ubicación de las estaciones de muestreo en Cabo Manglares - Distrito Nacional de Manejo Integrado Cabo Manglares Bajo Mira y Fronteras, costa Pacífica colombiana.
En laboratorio, fueron revisadas en su totalidad 128 muestras de zooplancton, siendo separadas, contadas e identificadas hasta el menor nivel taxonómico posible todas las larvas de peces. Para realizar la identificación taxonómica, se utilizaron las guías de Richards (2006) y Beltrán-Rios & Herrera (2000). Todo el material identificado fue depositado en la colección de referencia de Biología Marina de la Universidad del Valle (UNIVALLE: CIR-LV-UV).
La representatividad del muestreo se estableció como una proporción del valor esperado de riqueza para el área de estudio, definido a partir del estimador no paramétrico de riqueza Jackniffe de segundo orden (Heltshe & Forrester, 1983; Colwell & Coddington, 1994). Se comparó la abundancia de larvas de peces entre periodos utilizando una prueba no paramétrica de Mann Whitman (MW) y la riqueza de especies (Chao 1), dominancia (Simpson) y diversidad alfa (Shannon) entre periodos de muestreo mediante un análisis de comparación por remuestreo de Bootstrap, a partir de 10000 interacciones aleatorias.
RESULTADOS
En total fueron capturadas y procesadas 4947 larvas de peces durante los dos periodos de estudio. 153 unidades taxonómicas morfológicas de larvas de peces en la zona de estudio, pertenecientes a 43 familias, de las cuales 60 unidades se identificaron al nivel de especie, 45 a género y 48 a nivel de familia (tabla 1). La representatividad del muestreo fue mayor al 84 % (Jacknife 2 = 183 especies), encontrándose diferencias significativas en la riqueza de especies y la diversidad, pero no en la dominancia (tabla 2).
Tabla 1 Composición taxonómica del ensamblaje de larvas de peces en Cabo Manglares - Distrito Nacional de Manejo Integrado Cabo Manglares Bajo Mira y Fronteras durante diciembre 2016 y marzo 2017. Se presenta la abundancia media (AM) estandarizada por el volumen de agua filtrada y la abundancia relativa (AR) de cada identidad taxonómica morfológica definida.
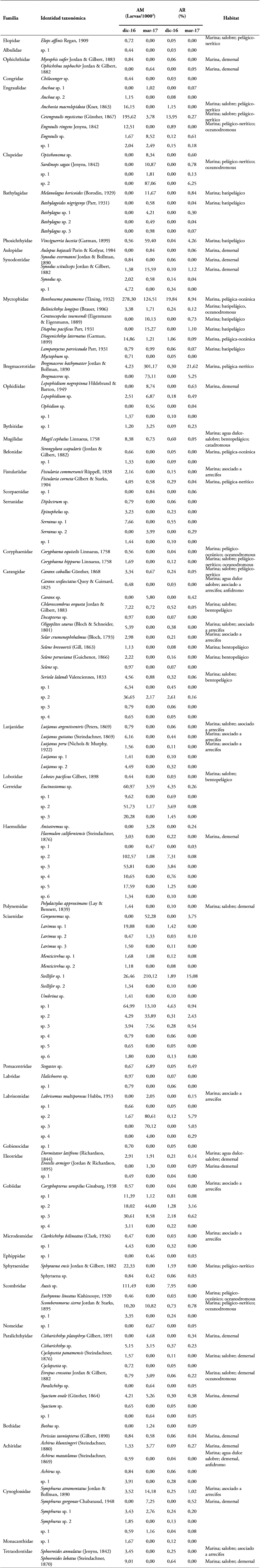
Tabla 2 Resultados de la comparación de la riqueza (Chao 1), dominancia (Simpson) y diversidad (Shannon) de larvas de peces entre diciembre 2016 y marzo de 2017 en Cabo Manglares - Distrito Nacional de Manejo Integrado Cabo Manglares Bajo Mira y Fronteras (valor-p establecido por permutación a partir de 1000 interacciones; n = 32 para cada periodo)

El 9,9 % de las larvas capturadas durante diciembre de 2016 fueron vitelinas con una abundancia media de 146 larvas vitelinas/1000 m3, mientras que durante marzo del 2017 solo el 2,7% fueron vitelinas con una abundancia media de 39 larvas vitelinas/1000 m3. La mediana de abundancia de larvas de peces considerando los dos periodos de estudio fue de 903 larvas/1000 m3. Aunque se registró una mayor abundancia de larvas de peces durante marzo de 2017 (mediana =973 larvas/1000 m3) que durante diciembre 2016 (mediana= 747 larvas/1000 m3), no se establecieron diferencias significativas entre los periodos de estudio (MW, p =0,943).
El 28,8 % de las especies identificadas fueron registradas en ambos periodos de muestreo, mientras que el 63 % de las especies identificadas solo fueron registradas durante diciembre de 2016 y el 43,6 % de las especies solo fueron registradas en marzo de 2017, siendo el porcentaje de similitud entre periodos del 40 % (índice de similitud de Bray Curtis).
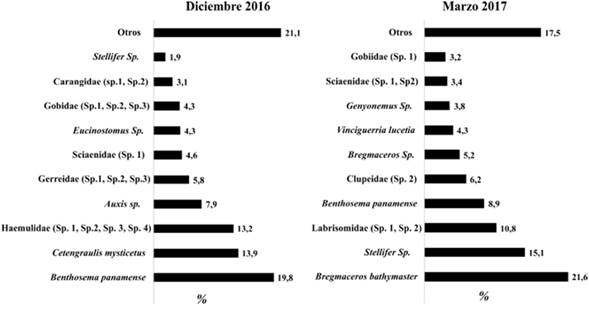
Durante diciembre de 2016 las familias más abundantes fueron Myctophidae, Engraulidae y Haemulidae, mientras que durante marzo de 2017 las familias más abundantes fueron Bregmacerotidae, Scianidae y Labrisomidae (figura 2). Las especies más abundantes durante diciembre de 2016 fueron Benthosema panamense, Cetengraulis mysticetus, Haemulidae y Auxis sp, mientras que durante marzo de 2017 las especies más abundantes fueron Bregmaceros bathymaster, Stelllifer sp. Labrisomidae y Benthosema panamense (figura 3).

Figura 2 Composición porcentual por familias del ensamblaje de larvas de peces en Cabo Manglares durante diciembre 2016 y marzo 2017.
DISCUSIÓN
El presente estudio reveló que el ensamblaje de larvas de peces en la zona de Cabo Manglares (DNMI Cabo Manglares, Bajo Mira y Frontera) es taxonómicamente diverso y exhibe una marcada diferencia estacional en términos de su composición, de manera similar a lo reportado para otros ambientes marino-costeros tropicales, toda vez que la composición de los ensamblajes de larvas de peces responde de manera muy estrecha a la variabilidad de las condiciones ambientales locales (Whitfield & Pattrick, 2015; Houde, 2008; León-Chavez et al., 2010; De Macedo-Soares et al., 2014; León-Chávez et al. 2015; Keskin & Pauly, 2019).
Establecer los factores que influencian las características estructurales de los ensamblajes de larvas de peces es complejo. La estructura espacial generalmente es determinada por las características hidrodinámicas de la región, como frentes térmicos o salinos o el patrón local de circulación superficial (Govoni, Hoss & Colby, 1989; Cowen, Hare & Fahay, 1993; Sabates & Olivar, 1996; Martínez-Aguilar et al., 2010; Calle-Bonilla et al., 2017), mientras que la estructura estacional generalmente está determinada por los procesos reproductivos y de desove de los adultos, procesos que son específicos para cada especie (Gray & Miskiewicz, 2000; Marancik, Clough & Hare, 2005).
Aunque la zona de Cabo Manglares sustenta el 24.5 % de la pesca artesanal que se genera en el Pacífico colombiano (AUNAP-UNIMAGDALENA, 2012), no hay referencias específicas sobre el ensamblaje de larvas de peces en esta localidad. Solo se puede derivar alguna información sobre este tema de los trabajos desarrollados por Beltrán-Leon & Rios (2000); Escarria et al. (2006) y Martínez-Aguilar et al. (2010), esfuerzos de investigación que abordaron el estudio de las larvas de peces en toda la cuenca Pacífica colombiana. Por lo tanto, el presente trabajo de investigación presenta información novedosa sobre la composición y estructura del ensamblaje de larvas de peces en la zona de Cabo Manglares, incluyendo información sobre la abundancia de estadios larvales de especies de interés pesquero artesanal para los habitantes de la región como el pargo (Lutjanidae: e.g. Lutjanus guttatus), Pelada (Scianidae), Picuda (Sphyraenidae: e.g. Sphyraena ensis), Sierra (Scombridae: e.g. Scomberomorus sierra), Corvina (Scianidae: e.g. Stellifer sp.) y Jurel (Carangidae: e.g. Caranx caballus), y la carduma (Engraulidae: e.g. Cetengraulis mysticetus) que es utilizada para carnada y procesamiento de la industria harinera (Ortiz & Lopez, 2015).
Los estudios taxonómicos sobre las larvas de peces son una aproximación adecuada para llenar los vacíos de información sobre una de las fases de desarrollo más crítica del componente íctico de una localidad. Además de proporcionar información que permite valorar la biodiversidad marina local (Warner et al., 2000; Hastings & Botsford, 2003), conocer la composición taxonómica de los ensamblajes de larvas de peces es el punto de partida para comprender las tendencias de variación espacio-temporales y relaciones ecológicas con el hábitat disponible por parte de los peces adultos (King & McFarlane, 2003; Costa, Possinghamy & Muelbert, 2015). El estudio aquí realizado, aunque es de pequeña escala espacial, proporcionan evidencia importante sobre la estructura del ictioplancton en ambientes del Pacífico colombiano, lo cual puede traer grandes beneficios al integrarse con estudios de dinámica poblacional de los estadios adultos y la caracterización de las condiciones oceanográficas propias de cada sitio de interés (Beldade, Borges & Gonçalves, 2006; Olivar et al., 2010). Además, proporciona información relevante para fortalecer las acciones de manejo del Distrito Nacional de Manejo Integrado Cabo Manglares, Bajo Mira y Frontera.

