










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAntipoda. Revista de Antropología y Arqueología
Print version ISSN 1900-5407
Antipod. Rev. Antropol. Arqueol. no.5 Bogotá July/Dec. 2007
EL CONOCIMIENTO ETÜLÓGICÜ COMO FANTASMA DE LA BIOLOGÍA Y SU IMPORTANCIA PARA LOS ESTUDIOS COMPARADOS1
Héctor Blass Lahitte2 María Paula Tujague3
2 Licenciado en Antropología, doctor en Ciencias Naturales y doctor en Psicología. Investigador principal CIC, Argentina. Director del instituto de investigación en Antropología Cognitiva, pinaco. Profesor titular de Teoría Antropológica y Etología. Miembro de la Academia Nacional de Ciencias de Córdoba y Buenos Aires. Profesor e investigador visitante en España. Profesor titular y jefe del Departamento Científico de Etnografía, Facultad de Ciencias Naturales y Museo, fcnym, Universidad Nacional de la Plata, Argentina lahitte@fcnym.unlp.edu.ar
3 Licenciada en Biología con orientación en Zoología. Becaria de iniciación y personal del Departamento Científico de Etnografía, Facultad de Ciencias Naturales y Museo, FCNyM, Universidad Nacional de la Plata, Argentina mpaulat@mdht.com.ar
RESUMEN
La distancia que separa al hombre de los demás animales no es tan abismal. Cuanto más aprendemos de ellos, más pertinentes son estos estudios para solucionar numerosos problemas humanos. El investigador confecciona un mapa de lo que supone un proceso conceptual interno, para registrar qué conjuntos de rasgos de conducta permiten describir índices y establecer relaciones. Nuestro objetivo es evidenciar la importancia del estudio de concordancias conductuales en mamíferos, como base para elaborar un marco teórico que permita interpretar la conducta del hombre y otros primates, analizando al espacio como contexto para entender modalidades de acoplamiento del organismo con su entorno.
PALABRAS CLAVE
Etología, biología, antropología, primates, espacio.
THE ETHOLOGIC KNOWLEDGE LIKE A BIOLOGY'S GHOST AND HIS IMPORTANCE FOR THE COMPARATIVE STUDIES
ABSTRACT
Distance between man and animal is not so deep. The more we learn from them, the more useful are the studies to solve human problems. To record behavioral features and use them to describe indexes and establish relations, observer makes a map about what is supposed to be the animal's conceptual process. Our goal is to prove the importance of studying behavioral concordances in mammals as a base to generate a theoretical framewok that allows the interpretation of human and other primate's behavior, taking into account the space as context to understand different ways organisms copes whit environment.
KEY WORDS
Ethology, Biology, Anthropology, Primates, Space.
FECHA DE RECEPCIÓN: ABRIL DE 2007 / FECHA DE ACEPTACIÓN: OCTUBRE DE 2007
En tanto que biólogo que estudia el comportamiento social de los mamíferos,
deploro profundamente la ausencia de un marco de pensamiento capaz de englobar
a la vez el comportamiento de los hombres y el de otros mamíferos.
En tanto que seres inteligentes que viven en la era del espacio,
deberíamos intentar remediar este estado de cosas...
(Michael Chance, 1978)
La práctica de la interdisciplinaridad supone la formación de nuevos especialistas y, desde luego, una reforma profunda de los sistemas educativos. Al mismo tiempo, se hace necesario iniciar a los jóvenes investigadores en reflexiones que permitan hacer progresar la teoría a partir de situaciones concretas.
Estudios recientes a propósito de fenómenos ecológicos planetarios modifican los ejes privilegiados y la organización de la investigación, tanto en ciencias naturales como sociales. Investigadores de uno y otro campo estudian la influencia del medio natural sobre los fenómenos sociales -incluso desde una perspectiva política- y el impacto de las actividades humanas sobre el medio natural. Es en este contexto donde el estudio del comportamiento se presenta como el nexo entre los unos y los otros.
La distancia que separa al hombre de los demás animales no es tan abismal como puede suponerse. Todos aceptamos que descendemos del "mono", pero ninguno de nosotros explica cuál es nuestro parentesco con él (Morin, 1980; Morin y Piatelli-Palmarini, 1983). Cuarenta años de trabajo de campo con primates han confirmado que somos una especie completamente distinta de cualquier otra. No existe un equivalente exacto al esquema humano. Pero en el reino animal no hay nada excepcional en esto. Cada especie es única (Ridley, 2004): "Cuanto más aprendemos de los animales y de los intrincados mecanismos de adaptación que la evolución ha producido, más pertinentes son estos estudios para la solución de algunos de los más complejos problemas humanos" (Hall, 1998).
Durante muchos años, zoólogos y antropólogos se abocaron al estudio de mensajes y modos de comunicación en mamíferos, en particular Gregory Bateson (1974; 1985), quien pautó la modalidad de mensajes en mamíferos pre-verbales. Los seres humanos no hacen evidente el componente afectivo en la comunicación. Éste queda encubierto por el lenguaje digital a pesar de que expresan, en muchos casos inconscientemente, el componente emocional con señales cinéticas y paralingüísticas. David Premack (1983) nos recuerda que este tipo de señales, que deberían haber entrado en decadencia en el caso de los seres humanos, distan mucho de ser anacrónicas.
Es frecuente que las conductas o coordinaciones conductuales se clasifiquen o tipifiquen según los resultados obtenidos en laboratorios experimentales o en reductos "a campo traviesa" -que están cercados o limitados por distancias mayores-. En realidad se deberían comparar ambos, no para hablar de la conducta en sí, sino para hablar de las variaciones que produce el tipo de medio en el organismo que da respuesta. Lo antedicho se hace evidente cuando zoólogos, etólogos y antropólogos analizan organizaciones jerárquicas como resultado de las más diversas coordinaciones conductuales. Se puede comprender la significación de lo que se observa sólo si se poseen los mecanismos interpretativos adecuados: registros de actos de conducta patronados, bases de datos integradas por rasgos de conducta mayores y menores, mapeo del territorio, inventario de necesidades, uso y acciones repetitivas, concordancia redundante, etcétera.
En una situación diferente, y a modo de ejemplo sobre estudios comparados, Gregory Bateson (1990) estudió las formas de interacción entre los nativos iatmules de Nueva Guinea. Demostró cómo parejas de varones habían practicado secuencias simétricas de conducta. Competían entre sí petulantemente en las ceremonias de iniciación de los novatos. Así también, las parejas de delfines nadan simétricamente y compiten en carreras saltando a veces al mismo tiempo... A la interacción entre hombres y mujeres iatmules, Bateson las calificó de complementarias -es decir, diferenciadas pero integradas-. Explicaba que el exhibicionismo de los varones empalmaba con la expectativa de las mujeres. Al comparar este patrón conductual con el de los delfines que estudió en Hawai, observó que interacciones en parejas de estos animales pautaban diversas relaciones complementarias: coito, amenazas, formas de propulsión, etcétera. La conclusión que de esto extrae Bateson puede sintetizarse como que, en ambos casos, se trata de interacciones tendientes a poner de manifiesto distribuciones jerárquicas entre individuos del mismo o diferente sexo, y/o formas de certificación social.
Este trabajo tiene por objetivo evidenciar la importancia del estudio de las concordancias conductuales en mamíferos, como base para la elaboración de un marco teórico que nos dé elementos para interpretar la conducta del hombre y de otros primates. Entre otros aspectos, analizaremos el espacio como contexto para entender las modalidades de acoplamiento del organismo con su entorno, y a la vez, subrayar la importancia de la ontogenia del individuo en dicho acoplamiento.
Como en todo trabajo con pretensiones zoosemióticas, transformaremos en argumentos los hechos de observación. Será desde la repetición y la redundancia desde donde hablaremos. La redundancia sólo es posible gracias a un mecanismo repetitivo de extracción de diferencias (Lahitte et ál., 1987).
RECURSOS COMUNICATIVOS EN PRIMATES NO HUMANOS Y HUMANOS
En el caso que nos ocupa, el de los primates en general, la interacción entre organismos -y de los organismos con su entorno- permite reconocer patrones comportamentales. A partir del comportamiento exhibido por el objeto de estudio, el investigador confecciona un mapa de lo que supone un proceso conceptual interno, para registrar qué conjuntos de rasgos de conducta permiten describir índices y establecer relaciones entre un estado y otro. Intentar hablar de significados para el animal pierde significación respecto de la investigación sobre lo que pueden hacer.
Desde luego, como plantea Premack (1983), cabe la pregunta sobre si es el hombre quien confecciona el mapa de su propia estructura cognitiva a partir de las analogías que descubre en el proceso de interacción con los primates, o bien, si lo estudiado y/o cartografiado es la estructura cognitiva de los animales en cuestión.
Heinz von Foerster (1988) propuso una alternativa teórica importante que suponía la posibilidad de generar un marco experimental en el que se hiciera evidente si los primates en cuestión son capaces de distinguir la simetría, la transitividad o la reflexividad como índices de equivalencia, o si por lo contrario, reconocen índices del concepto de diferencia. De hacerlo, expresarían su capacidad para operaciones lógicas y el dilema quedaría resuelto: un mapa -map- puede ser mapeado -mapped-. Esto, a su vez, permitirá al etólogo analizar cómo una señal se vuelve signo, lo que no significa abordar dichos acontecimientos desde la perspectiva de la significación sino desde la de la comunicación. En otras palabras, describir qué y cómo comunican, y no interpretar cuál es el significado de sus acciones.
Para poder hablar -argumentar sobre los significados que "el otro" asigna a la realidad con la que interactúa-, deberíamos atribuir los mismos valores a los objetos del mundo y, desde luego, no hay forma de probar dicho postulado. Es muy probable que la comunicación "entre" primates sea analógico-afectiva y no simbólica, y la traducción de una a otra entrañe un cierto número de problemas. La comunicación analógico-afectiva está sujeta a la concordancia de preferencias, mientras que la simbólica no.
Si bien es difícil persuadir a quienes suponen que el significado es evidente -se trate de primates, delfines o perros-, conviene recordar, a profanos e iniciados, que si no podemos extraer una matriz con la que los organismos estudiados ordenan los acontecimientos del medio, resulta desde todo punto de vista improbable hablar de comunicación "simbólica".
David Premack (1977) nos recuerda el experimento de Menzel con chimpancés en cautiverio: se observa, describe y registra qué y cómo comunican, luego se anulan las escalas de preferencia -tipo de ámbito, cosa intercambiada, hora, etcétera- y luego se analiza si a pesar de ello pueden o siguen comunicándose, es decir, si tienen comunicación simbólica. Lo probado es que una vez destruida la concordancia de preferencias -estado a- no pueden, hasta reconstruirla, acceder a la comunicación.
Sin duda, los trabajos de Michael Chance (Chance y Jolly, 1980) en Bermingham con la colonia de Macaca fascicularis constituye un gran aporte al desarrollo teórico de las próximas generaciones de etólogos. Chance realizó un aporte teórico fundamental para entender las condiciones y mecanismos que subyacen a toda organización grupal -¿social?-, tanto en el hombre como en otros mamíferos, en particular primates. Para llevar a cabo sus investigaciones registró el comportamiento de monos en cautiverio y en estado salvaje, llevando a cabo distintas formas de control experimental. En sus últimos trabajos, Chance describe patrones de conducta como conjuntos de rasgos que permiten distinguir comportamientos hedónicos y agonísticos, conjuntamente con las estructuras atencionales que dan coherencia al grupo. En el comportamiento agonístico todas las interacciones están destinadas a concluir con la interacción en sí, es decir, son modalidades de evitación. En general, el comportamiento agonístico evita el ataque evitando el contacto, mientras que la activación hedónica evita el ataque por el control del contacto.
En sus textos, Chance explica de qué manera los componentes de un grupo se asocian según las formas de "atención" de que dispongan. Tipifica dos modos centrípetos: a) el asociado al rango en babuinos y macacos, y b) el referido al líder en monos antropomorfos.
En los monos caí o capuchinos, Cebus apella, encontramos que el ejemplo se ajusta al modo b, donde la dirección de desplazamiento del individuo -macho- al que se adjudica el rasgo de "dominante" será la que adoptará tarde o temprano la totalidad del grupo. Como uno de nosotros pudo observar en sus trabajos en campo, junto al grupo de investigación del Dr. Charles Janson en la provincia de Misiones, Argentina, el cambio en la ruta de desplazamiento de este individuo lleva a todo el grupo a modificar el suyo, para escoger, por ejemplo, un sitio dormidero para pasar la noche. En una ocasión en la que esto ocurrió, media hora después del cambio de dirección del macho dominante y de su cambio en la elección del sitio donde pasar la noche, seguían arribando al nuevo sitio individuos que habían mantenido la dirección inicial.
Es interesante recalcar que, por debajo de este "líder", pareciera existir un modo centrípeto de tipo a, que vale tanto para machos como hembras del grupo. Los mismos se ubicarán en una escalera jerárquica que quedará en evidencia para el observador por medio de la descripción de determinados rasgos y atributos comportamentales. En la actualidad, las jerarquías de dominancia para los grupos de primates son establecidas por los investigadores a través del análisis de las interacciones diádicas en las peleas. Nos parece interesante el enfoque de Chance, porque ofrece una posible matriz de análisis para eliminar la relatividad de la cuestión.
El objetivo privilegiado de los experimentos de Chance fue tipificar las focalizaciones atencionales -de un miembro cuyo menor gesto redistribuye posturas y actividades de los demás- y los universales de la atención.
Chance concluye que un buen insight del observador con un referente empírico, permitirá comprender mejor las bases constitutivas de la sociedad humana. En las conclusiones dice:
En una sociedad humana en la que el comportamiento social se ha desmoronado, los individuos prestan atención excesiva a ciertas personalidades. Forman una especie de séquito detrás del personaje carismático (...) propensión que sin duda tiene que ver con nuestra condición de primates (Chance y Jolly, 1980).
Por último, consideramos que es importante aclarar que no existe una epistemología del fantasma. Es imposible presuponer que el observador en campo no interfiere en el comportamiento de los objetos de estudio. En las observaciones en campo es frecuente que, por ejemplo, frente a una interacción agresiva entre dos o más individuos, el individuo al que se dirigen las pautas agresivas realice los mismos comportamientos pero dirigidos al observador que se encuentra allí en ese momento, aun cuando los objetos de estudio no escapan a la presencia del investigador, tolerando la misma a lo largo del día.
COMPORTAMIENTO Y ESPACIO
El espacio es una variable que, en general, es dejada de lado en los estudios de comportamiento animal, particularmente en aquellos referidos al comportamiento humano (Campos-de-Carvalho et ál., 2004). Es difícil concebir algo que no esté ubicado en tiempo y espacio (Zubrow y Daly, 1998). Las nociones de tiempo, espacio y causalidad, según nuestras percepciones, orientan la representación que tenemos del mundo y de nosotros mismos (Jacob, 1998).
El espacio percibido no es homogéneo. En el uso que cada individuo hace del mismo, el observador podrá realizar un mapeo e inferir a partir de ello la estructura espacial interna o configuración del organismo en cuestión. A su vez, ese mapa evidencia la propia estructura cognitiva del observador, ya que dos observadores en un mismo contexto distinguirán cosas diferentes. El observador construye configuraciones de lo que percibe, siendo siempre el punto de vista humano del quehacer animal (Lahitte et ál., 2005).
Es importante recordar que lo que vemos son esquemas de acción, no mapas o modelos internos (Lahitte et ál, 2003: 166). El dato no es el hecho en el mismo sentido que afirmamos que el mapa no es el territorio sino su representación. Los hechos -territorio- no aparecen nunca en el modelo -mapa-(Lahitte et ál., 1987). El modelo es confeccionado por el observador.
En los mamíferos superiores, el sentido del espacio es una síntesis de la entrada de datos sensoriales de muchos tipos. Al mismo tiempo, en el hombre, cada uno de ellos es modelado y configurado por la cultura. Cada individuo tiene un modo diferente de sentir el espacio. En la historia de los estudios de comportamiento espacial humano fue, sin dudas, Edward Hall, quien rompió con las estructuras clásicas al desarrollar la proxémica: "Se entiende por proxémica a la descripción de las observaciones y teorías interrelacionadas acerca del empleo del espacio por parte del hombre" (Hall, 1998: 125). Las normas proxémicas desempeñan un papel comparable al de los movimientos expresivos en animales inferiores, es decir, consolidan el grupo al mismo tiempo que lo aíslan de los demás, reforzando por una parte la identidad intragrupal y dificultando, por la otra, la comunicación intergrupal.
Las personas criadas en culturas diferentes viven en mundos sensoriales distintos. En sus estudios, Hall demostró que en los humanos existen importantes diferencias culturales en el tratamiento del espacio. En el empleo del espacio interior, por ejemplo, los japoneses dejan despejados los rincones de las piezas porque todo sucede en el medio. Los europeos tienen tendencia a llenar los rincones poniendo los muebles cerca de las paredes o pegados a ellas. En la cultura norteamericana, el espacio necesario es el que se requiere estrictamente para desempeñar una tarea. Todo lo que pase de la necesidad mínima suele considerarse superfluo. En el Oeste, el hombre no percibe los objetos sino el espacio que hay entre ellos. En el Japón, los espacios se perciben, denominan y reverencian bajo el nombre de ma o hueco intermedio. Cada cultura posee una concepción del espacio totalmente diferente.
El desarrollo de habilidades cognitivas vinculadas al espacio presenta un marcado dimorfismo sexual. Los hombres superan a las mujeres en el desarrollo de habilidades espaciales. Un dimorfismo semejante ha sido observado en monos rhesus, Macaca mulatta, donde los machos jóvenes superan a las hembras jóvenes. Esta "superioridad"' de los machos desaparece con la edad, dando como resultado que machos adultos no superen a las hembras adultas en este aspecto (Driscoll y Sutherland, 2005). Según observadores humanos, los machos de mamífero, en general, tienen mayores habilidades para manejarse en el espacio que las hembras, lo que se correlaciona con una tendencia en estos a que prevalezca la preferencia de señales de configuración del espacio para moverse en él, mientras que las hembras prefieren señales de color o de forma. Acorde a esto, los machos presentan un mayor desarrollo del tamaño del hipocampo (Hodgson y Healey, 2005).
En un estudio sobre comportamiento lúdico en niños y niñas norteamericanos de diez a doce años de edad (Erikson, 1980), se encontró que los mismos utilizaban el espacio para el juego de un modo muy diferente. Al comparar las construcciones en el espacio tridimensional que realizaban al jugar, Erikson concluyó que determinadas configuraciones se daban con notable frecuencia en un sexo y no en el otro. La escena construida por una niña representaba el interior de una casa, con personas y animales en su mayoría dentro de tal recinto y en posición estática -sentados o de pie-. Las escenas construidas por los niños eran casas con muros o fachadas detalladas -enfatizando el exterior de la vivienda-, y la mayoría de las personas y animales se situaban fuera de recintos o edificios, con más vehículos y animales que se movían por las calles.
Numerosos investigadores consideran que la posición de cada individuo durante los desplazamientos en una tropa de primates, se relaciona estrechamente con la ubicación jerárquica de los mismos en el grupo, y por qué no, con una historia evolutiva de configuraciones internas o externas del espacio. Erikson pudo observar una analogía entre los comportamientos de construcción de juguetes y la morfología de grupo de los babuinos:
En sus desplazamientos diarios, el centro del grupo está ocupado -a modo de un espacio interno- por las hembras grávidas o que transportan a sus crías, mientras que en los alrededores se ubican los machos. Sean cuales fueren las diferencias morfológicas entre babuinos machos y hembras en cuanto a estructuras óseas, posturas y comportamientos, parecieran estar adaptados a sus respectivas tareas de albergar y defender los círculos concéntricos desde la matriz procreadora hasta los límites del territorio defendible (Erikson, 1908).
Algo semejante puede observarse en los monos caí, que muestran una marcada estructura espacial durante la búsqueda y localización del alimento (Di Bitetti y Janson, 2001). Los adultos dominantes se ubican en la zona anterior y central, los infantes y juveniles en el centro y los adultos subordinados en las periferias (Fragaszy et ál., 2004). Creemos que es bueno aclarar que los estudios de Erikson se limitan a la sociedad californiana, lo que no es un detalle a la hora de generalizar, como ya nos han demostrado los estudios de Hall sobre numerosas culturas.
En el libro de Edward Hall (1998), y a modo de ejemplo de lo determinante que es la configuración espacial en el comportamiento, se comenta la experiencia de Humphrey Osmond en una sala modelo de geriatría femenina en el Centro de Salud e Investigación en Saskatchewan, Canadá. La sala era nueva y espaciosa pero, cuanto más estaban allí las pacientes, menos hablaban entre ellas. Poco a poco se iban pareciendo a los muebles, pegados a las paredes a intervalos regulares entre las camas. Advirtiendo que el espacio era más soció-fugo -es decir que tiende a mantener apartadas a las personas- que sociópeto -es decir que tiende a reunir a las personas-, Osmond encargó a un joven y perceptivo psicólogo llamado Robert Sommer para que estudiara la cuestión.
En cincuenta sesiones de observación en la cafetería del mismo hospital, Sommer contó el número de conversaciones que ocurrían en las mesas rectangulares de la misma, según las ubicaciones de los individuos. Las conversaciones en un rincón de la mesa eran el doble de frecuentes que las que se daban entre dos personas sentadas de un mismo lateral, y a su vez, éstas eran tres veces más frecuentes que las que ocurrían entre personas sentadas en lados opuestos a lo ancho de la mesa. En lados opuestos, a lo largo, no se observó conversación alguna.
Realizando cambios en la ubicación y el tipo de mobiliario en la sala de geriatría femenina, Sommer y Osmond lograron duplicar el número de conversaciones de las pacientes en la sala.
Además de las diferencias de sexo en el desarrollo de habilidades espaciales, existen rasgos individuales en la respuesta frente al espacio que no pueden dejarse de lado. En un total de veinte horas de observaciones preliminares sobre dos individuos de mono caí en condiciones de semicautiverio, en una isla artificial del Jardín Zoológico y Botánico de La Plata, Buenos Aires, concluimos que la respuesta de comportamiento frente al espacio disponible en el encierro es individual y depende de la historia de cada organismo. Así, se encontraron importantes diferencias en el grado de desarrollo de comportamientos estereotipados en los dos individuos, a pesar de que ambos llevan el mismo período de tiempo en ese ambiente pero sus historias individuales eran completamente distintas -datos proporcionados por el personal del zoológico-. De un total de novecientos ochenta y seis eventos de estereotipos registrados, un 99,18 por ciento correspondía al individuo 1 -macho-, mientras que el 0.81 por ciento restante se registró para el individuo 2 -hembra- (Gráfico 1).
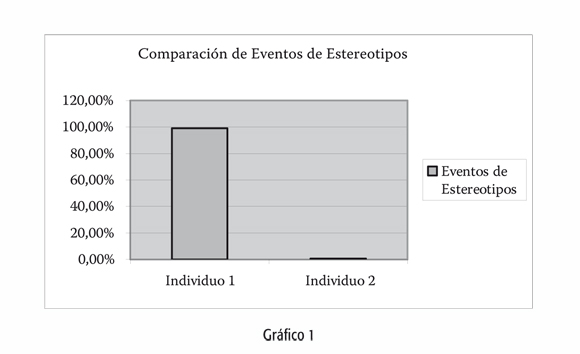
Los estereotipos de desplazamiento son considerados, por regla general, como un resultado anormal del encierro (Philbin, 2007; Roder y Timmermans, 2002). Entendemos por comportamientos estereotipados a aquellas secuencias altamente repetitivas y uniformes, que carecen de función aparente (Wemelsfelder, 1993). Frente a la baja incidencia de registro de estos comportamientos en el individuo 2, cabe la pregunta de si es correcto asignarles esta categoría. Creemos que el hecho de no haber observado ninguno de los dos comportamientos descritos por nosotros como estereotipos de encierro a lo largo de los meses de observación de la especie en libertad, es un indicador de su anormalidad. Por otra parte, los mismos comportamientos han sido reportados por otros investigadores en la misma especie en cautiverio (Pavé, comunicación personal; Polizzi, comunicación personal). Concluimos que un espacio reducido no es por sí solo ni obligatoriamente un disparador de conductas anormales.
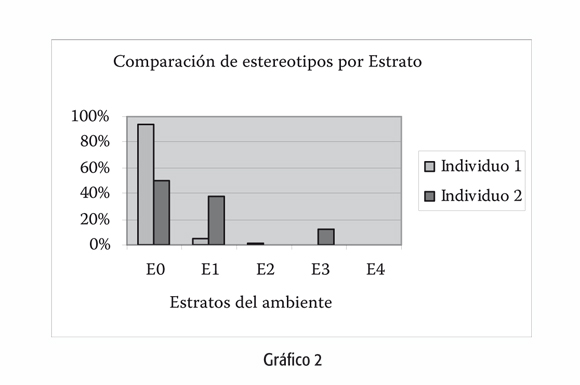
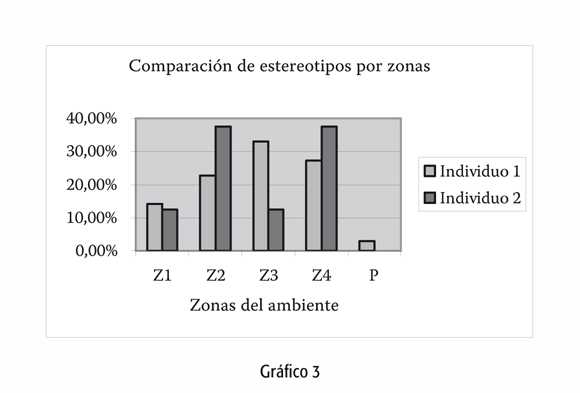
A la vez, existen rasgos espaciales asociados a la especie en sí. En el mapeo que realizamos en el mismo ambiente, el uso del espacio horizontal -diferentes zonas del ambiente, gráfico 2- resulta ser más homogéneo que el vertical -estratos del ambiente, gráfico 3-, acorde a una especie arborícola, a pesar del encierro. No cabe duda que un organismo que se mueve sobre el nivel del suelo, en los estratos de los árboles, percibe el espacio de manera diferente a uno que sólo se desplaza por el piso. La visibilidad es diferente, la superficie de apoyo es distinta. Llamativamente nuestras conclusiones muestran que la mayor parte de los comportamientos anormales se registran al nivel del suelo, el cual es el estrato menos utilizado por la especie en libertad.
Las diferentes maneras de organizar el espacio generan una amplia diversidad de comportamientos sociales. El espacio nunca es neutral: la presencia o ausencia de algún elemento, y su organización, siempre transmite un mensaje (Campos-de-Carvalho et ál., 2004). Cada organismo, en un espacio dado, mapeará el mismo según su estructura cognitiva, y operará en él a partir de ese mapeo. El espacio mapeado incluirá, por supuesto, otros organismos.
Campos-de-Carvalho (2004), en su estudio sobre respuestas de comportamiento de niños en jardines de infantes, midió en función de distintos arreglos espaciales las diferentes actividades que los niños llevaban a cabo en forma individual, entre sí y con los adultos que estaban cuidándolos.
Encontró que en espacios más vacíos -sin mobiliario-, los niños se mantenían en las cercanías del adulto presente. A diferencia de esto, en un arreglo espacial que presentara zonas circunscriptas por muebles, los niños utilizaban menos las zonas alrededor del adulto, y pasaban más tiempo dentro de las áreas semicerradas formadas por el mobiliario, al mismo tiempo que interactuaban más entre sí y realizaban menos actividades en aislamiento.
De lo anteriormente expuesto se desprende que los estudios comparados con animales nos ayudan a comprender que el entorno influye en las necesidades espaciales de cualquier organismo vivo. En los animales podemos observar, a diferencia de lo que ocurre con humanos, la dirección, el ritmo y la amplitud de la modificación de comportamiento que siguen a los cambios en el espacio que tienen disponible los individuos (Hall, 1998). Limitando nuestras observaciones al modo que tienen los animales de situarse y utilizar el ámbito, es posible identificar contextos y modalidades de acción traducibles a lo humano. Una prueba significativa de que las personas criadas en culturas diferentes viven en mundos perceptuales distintos está en su modo de orientarse en el espacio, trasladarse y operar en él. Al desplazarse, consciente o inconscientemente, el hombre mapea sus trayectos y sistematiza sus desplazamientos. No se trata sólo de lo que los hombres configuramos al mirar sino de lo que hacemos al estar. El espacio se transforma en un disparador de acciones. De la misma manera, el modo en que, como etólogos, designamos lo que vemos tiene que ver con el modo en que identificamos lo que nuestro objeto de estudio hace.
A MODO DE GLOSARIO
Para evitar la ambigüedad terminológica que, inevitablemente, introduce dificultades epistemológicas, teóricas y metodológicas, presentaremos definiciones breves de algunos términos relevantes para el tema tratado en este artículo.
Entorno: En la ecología clásica se define al entorno como la materia-energía que rodea al organismo y con el cual éste interactúa. Así definido, el entorno implica una instancia ajena al organismo, con un carácter de cosa en sí misma debido al estatus bioenergético que se le adjudica. En este sentido se emplean como sinónimos los términos medio y ambiente.
Para tratar de evitar esta disociación preferimos definir al entorno como todo aquello que no queda especificado como parte del organismo por su propia organización, incluidos otros organismos (Lahitte et ál., 1987).
Medio como sinónimo de entorno: El medio de cualquier sistema es todo aquello que no queda determinado como parte de él por su organización, y que puede interactuar con él como unidad.
Entonces podemos afirmar que todo lo que ocurre en un organismo surge en él y está determinado por su estructura, y es esta estructura la que determina las configuraciones estructurales del medio con las que el organismo se puede encontrar en sus interacciones con él. Las interacciones con el medio sólo pueden disparar en el organismo cambios estructurales determinados en él (Maturana, 2003).
Contexto: Conjunto de alternativas o variables entre las cuales un organismo puede efectuar su próxima elección (Lahitte y Ortiz Oria, 2005). El contexto es el espacio -no necesariamente físico- dentro del cual se verifican los componentes y sus relaciones de modo tal que determine las fronteras -no necesariamente físicas- de la unidad en tanto tal. El hecho de hacer prescindible la consideración de espacios y fronteras físicas radica en que las dimensiones de contexto están dadas por las relaciones interpartes -estructura- que especifican la unidad particular. El contexto es un espacio relacional (Lahitte et ál., 1987).
Espacio: Continente de todos los objetos sensibles que coexisten /Capacidad de terreno, sitio o lugar /Transcurso de tiempo /Parte de este continente que ocupa cada objeto sensible y también parte no ocupada entre varios objetos sensibles (Diccionario Enciclopédico Abreviado Espasa-Calpe, 1957).
Continente de todos los cuerpos sensibles que coexisten. El espacio puede contener materia, pero también existe sin ella. En filosofía, el problema del espacio ha sido objeto de consideración. Para Platón, el espacio es el receptáculo de las cosas creadas, carece de figura y su propiedad fundamental es la de ser llenado; Aristóteles lo concibe como algo sin lo cual no es posible concebir las cosas, algo que "emana" de ellas y es sinónimo de lugar. La teoría de la relatividad ha cambiado radicalmente las antiguas concepciones científicas de la noción de espacio, relacionándolo íntimamente con la de tiempo (Monitor Enciclopedia, 1971).
En nuestra concepción, entendemos al espacio físico como una variable dentro del contexto en el que se encuentra el organismo o individuo, que participa en la determinación del sistema organismo-entorno, y a la vez del sistema observador-entorno. La forma en que esa variable influye en el organismo queda determinada en la relación. La configuración espacial disponible activará en el organismo sólo aquello determinado en la estructura del organismo como tal. Y en consecuencia, éste operará sobre ese espacio en función de esa relación.
Gráficos 2 y 3:
Estrato 0: Nivel del suelo.
Estrato 1: Por encima del suelo hasta la altura de dos metros. Estrato 2: Entre dos y seis metros de altura.
Estrato 3: Entre seis y diez metros de altura, corresponde al límite superior de los árboles.
Estrato 4: Copas finales del eucalipto y las palmeras -superior a diez y hasta veinte metros de altura-.
z1 a z4: Zonas o cuadrantes en las que se dividió la isla para agilizar el registro.
p: Península. Porción de tierra delgada en un extremo de la isla. Poco uti+lizada por los individuos.
PAUTAS DE COMPORTAMIENTO ESTEREOTIPADO
-TOMADAS DE NUESTRO ETOGRAMO PARCIAL DE LA ESPECIE DE ESTUDIO-
Giro de cabeza:Durante el desplazamiento, el individuo realiza un movimiento de izquierda a derecha con la cabeza, llevando ésta primero hacia atrás y a la izquierda, luego hacia adelante y hacia la derecha.
Giro en 360°: Durante el desplazamiento, el individuo se detiene y rota su cuerpo de forma circular en el mismo lugar, quedando en la misma posición que antes de moverse, para luego continuar la marcha. El único miembro que se despega del suelo es el anterior izquierdo, en el momento de rotar. Esta pauta siempre es precedida por el giro de cabeza.
Comentarios
1 Agradecimientos para la producción de este artículo al Dr. Charles Janson y a la bióloga María Celia Baldovino por brindarnos la posibilidad de participar en su grupo de trabajo. A Alejo Castillo. Al Jardín Zoológico y Botánico de la ciudad de La Plata.
REFERENCIAS
Bateson, Gregory 1974 "Observations of a Cetacean Community", en Mindin the Waters, New York, Scribners. [ Links ]
Bateson, Gregory 1985 Pasos hacia una ecología de la mente, Buenos Aires, Editorial Lohlé. [ Links ]
Bateson, Gregory 1990 Naven. Un ceremonial Iatmul, España, Editorial Júcar Universidad. [ Links ]
Campos-de-Carvalho, M. I., R. Martínez, S. Morato de Carvalho, B. D. Resende, P. Izar, E. B. Ottoni, A. Carvalho y M. I. Pedrosa 2004 Dossier: "Space and Behaviour, a Comparative Perspective", en Revista de Etología, Vol. 6, No. 1, pp. 39-40. [ Links ]
Campos-de-carvalho, Mara 2004 "Use of Space by Children in Day Care Centers", en Revista de Etología, Vol. 6, No. 1, pp. 41-48. [ Links ]
Chance, Michael 1978 L'unité de l'homme, France, Ed. D'Seuil. [ Links ]
Chance, M. y C. Jolly 1980 Social Groups of Monkeys, Apes and Man, New York, Dutton. [ Links ]
Di Bitetti, M. S. y c. H. Janson 2001 "Social Foraging and the Finder's Share in Capuchin Monkeys, Cebus apella", en Animal Behavior, No. 62, pp. 47-56. [ Links ]
Driscoll, i. y R. J. Sutherland 2005 "The Aging Hippocampus: Navigation between Rat and Human Experiments", en Reviews in the Neurosciences, No. 16, pp. 87-121. [ Links ]
Erikson, Erik 1908 "Femineidad y espacio interior", en Identidad, juventud y crisis, cap. vii, Madrid, Taurus Ediciones. [ Links ]
Fragaszy, D. M, E. Visalberghi y L. M. Fedigan 2004 The Complete Capuchin: The Biology of the Genus Cebus, United Kingdom, Cambridge University Press. [ Links ]
Hall, Edward 1998 La dimensión oculta, 18a edición, España, Siglo XXI Editores. [ Links ]
Hodgson, Z. G. y S. D. Healy 2005 "Preference for Spatial Cues in a Non-Storing Songbird Species", en Animal Cognitio, No. 8, pp. 211-214. [ Links ]
Jacob, Francois 1998 El ratón, la mosca y el hombre, Barcelona, Drakontos. [ Links ]
Lahitte, H. B., J. A. Hurrell y A. R. Malpartida 1987 Relaciones. De la ecología de las ideas a la idea de ecología, Buenos Aires, Mako Editora. [ Links ]
Lahitte, H. B., H. R. Ferrari, V. Ortiz Oria y L. c. Lázaro 2003 Sobre la conducta como articulación individuo-entorno, Vol. III, 2a ed., Buenos Aires, Kliczkowsky Ediciones. [ Links ]
Lahitte, H. B. y V. Ortiz Oria 2005 El otro: antropología del sujeto, Buenos Aires, Editorial Nobuko. [ Links ]
Lahitte, H. B., R. H. Ferrari y L. c. Lázaro 2005 Etogramática: teoría y práctica de la descripción en ciencias del comportamiento, Buenos Aires, Nobuko. [ Links ]
Maturana, H. R. 2003 Desde la biología a la psicología, Buenos Aires, Lumen. [ Links ]
Morin, Edgar 1980 La méthode, France, Seuil. [ Links ]
Morin, E. y M. Piatelli-Palmarini 1983 El primate y el hombre, España, Editorial Argos Vergara. [ Links ]
Philbin, Nora 2007 "Towards an Understanding of Stereotypic Behaviour in Laboratory Macaques", disponible en www.pim.primate.wisc.edu/aboutp/behabior/index.html. [ Links ]
Premack, David 1977 Intelligence in Ape and Man, New York, Halsted Press for John Willey and Sons. [ Links ]
Premack, David 1983 "Capacidad de representación y accesibilidad del saber: El caso de los chimpancés", en N. Chomsky y J. Piaget. (eds.), Teorías del lenguaje y teorías del aprendizaje, pp. 254-285, Barcelona, Grijalbo. [ Links ]
Ridley, Matt 2004 Qué nos hace humanos, Madrid, Taurus. [ Links ]
Roder, E. L. y P. J. A. Timmermans 2002 "Housing and Care of Monkeys and Apes in Laboratories: Adaptations allowing Essencial Species-Specific Behaviour", en Laboratory Animals, No. 36, pp. 221-242. [ Links ]
Von Foerster, Heing 1988 "Construyendo una realidad", en Watzlawick et ál. (eds.), La realidad inventada, Buenos Aires, Gedisa. [ Links ]
Wemelsfelder, F. 1993 "The Concept of Boredom and its Relationship to Stereotyped Behavior", en A. B. Lawrence y J. Rushen (eds.), Stereotypic Behavior: Fundamentals and Applications to Animal Welfare, Wallingford, CAB International. [ Links ]
Zubrow, E. B. W. y P. T. Daly 1998: "Symbolic Behaviour. The Origin of a Spatial Perspective", chapter 13, en Colin Renfrew y Chris Scarre (eds.), Cognition and Material Culture: The Archaeology of Symbolic Storage, Cambridge, McDonald Institute for Archaeological Research, University of Cambridge. [ Links ]
OTRAS FUENTES
1957 Diccionario Enciclopédico Abreviado Espasa-Calpe, 7a ed., España. [ Links ]
1971 Monitor Enciclopedia, España, Salvat. [ Links ]

